Tilbage Klinik Klinisk Neurofysiologi Support. El Paso, TX. Kiropraktor, dr. Alexander Jimenez diskuterer klinisk neurofysiologi. Dr. Jimenez vil udforske den kliniske betydning og funktionelle aktiviteter af perifere nervefibre, rygmarven, hjernestammen og hjernen i forbindelse med viscerale og muskuloskeletale lidelser. Patienterne vil opnå en avanceret forståelse af smertens anatomi, genetik, biokemi og fysiologi i relation til forskellige kliniske syndromer. Ernæringsbiokemi relateret til nociception og smerte vil blive indarbejdet. Og implementeringen af denne information i terapiprogrammer vil blive fremhævet.
Vores team sætter en stor ære i at give vores familier og tilskadekomne patienter kun dokumenterede behandlingsprotokoller. Ved at undervise i komplet holistisk velvære som en livsstil, ændrer vi også ikke kun vores patienters liv, men også deres familier. Vi gør dette, så vi kan nå ud til lige så mange El Pasoans, som har brug for os, uanset overkommelighedsproblemer. For svar på eventuelle spørgsmål, du måtte have, ring venligst til Dr. Jimenez på 915-850-0900.
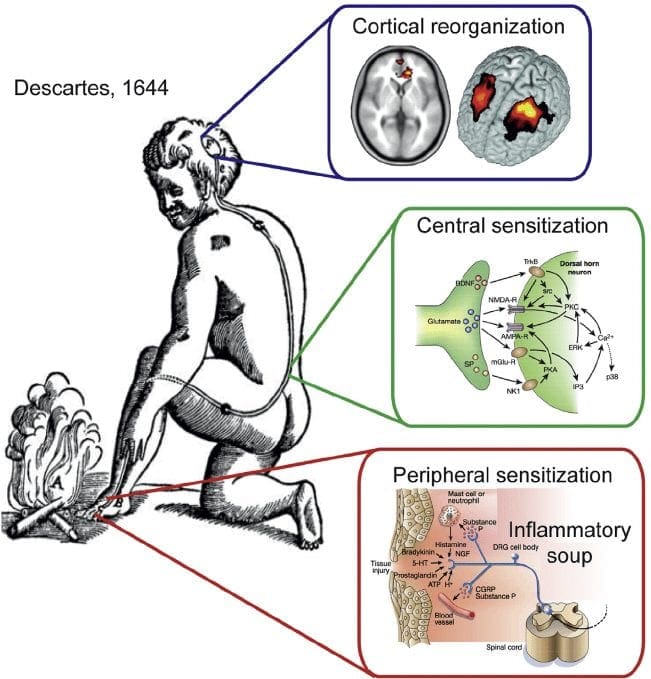
Neuropatisk smerte er en kompleks, kronisk smerte tilstand, som generelt ledsages af blødt vævsskade. Neuropatisk smerte er almindelig i klinisk praksis og udgør også en udfordring for både patienter og klinikere. Med neuropatisk smerte kan selve nervefibrene være beskadiget, dysfunktionelt eller skadet. Neuropatisk smerte er et resultat af skader fra traumer eller sygdomme til perifert eller centralnervesystem, hvor læsionen kan forekomme på et hvilket som helst sted. Som følge heraf kan disse beskadigede nervefibre sende ukorrekte signaler til andre smertecentre. Effekten af en nervefiberskade består af en ændring i neurale funktion, både i skadeområdet og også omkring skaden. Kliniske tegn på neuropatisk smerte indbefatter normalt sensoriske fænomener, såsom spontan smerte, paræstesier og hyperalgesi.
Neuropatisk smerte, som defineret af International Association of Pain Study eller IASP, er smerte initieret eller forårsaget af en primær læsion eller dysfunktion i nervesystemet. Det kan skyldes skader overalt langs neuraxerne: perifert nervesystem, spinal eller supraspinal nervesystem. Træk, der adskiller neuropatisk smerte fra andre former for smerte, omfatter smerte og sensoriske tegn, der varer ud over genopretningsperioden. Det er karakteriseret ved mennesker ved spontan smerte, allodyni eller oplevelsen af ikke-skadelig stimulering som smertefuldt og kausalgia eller vedvarende brændende smerte. Spontan smerte omfatter fornemmelser af "stifter og nåle", brændende, skydning, stabbing og paroxysmal smerte eller elektrisk stød som smerte, der ofte er forbundet med dysestesier og paræstesier. Disse følelser ændrer ikke blot patientens sensoriske apparat, men også patientens trivsel, humør, opmærksomhed og tænkning. Neuropatisk smerte består af både "negative" symptomer, såsom sensorisk tab og prikkende fornemmelser, og "positive" symptomer, såsom paræstesier, spontan smerte og øget følelse af smerte.
Betingelser, der ofte er relateret til neuropatisk smerte, kan klassificeres i to hovedgrupper: smerter på grund af beskadigelse i centralnervesystemet og smerte på grund af skade på det perifere nervesystem. Corticale og sub-kortikale slagtilfælde, traumatiske rygmarvsskader, syringo-myelia og syringobulbia, trigeminale og glossopharyngeale neuralgier, neoplastiske og andre pladskrævende læsioner er kliniske tilstande, der hører til den førstnævnte gruppe. Nerve kompression eller fastklemning neuropatier, iskæmisk neuropati, perifere polyneuropatier, plexopatier, nerveroden kompression, efter amputering stump og fantomsmerter, postherpetisk neuralgi og cancer-relaterede neuropatier er kliniske betingelser, der hører til den sidstnævnte gruppe.
Patofysiologi af neuropatisk smerte
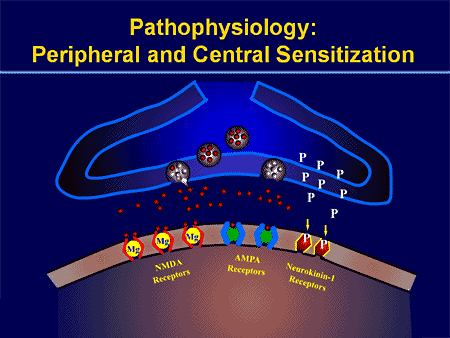
De patofysiologiske processer og begreber underliggende neuropatisk smerte er multiple. Forud for at dække disse processer er en gennemgang af almindelige smerte kredsløb kritisk. Regelmæssige smerte kredsløb involverer aktivering af en nociceptor, også kendt som smerte receptor, som reaktion på en smertefuld stimulering. En bølge af depolarisering leveres til de første ordens neuroner sammen med natriumhastning ind via natriumkanaler og udstødning af kalium. Neuroner slutter i hjernestammen i trigeminukernen eller i rygmarven på rygmarven. Det er her, hvor tegnet åbner spændingsgatede kalciumkanaler i den pre-synaptiske terminal, hvilket gør det muligt for calcium at komme ind. Calcium tillader glutamat, en excitatorisk neurotransmitter, at frigives til det synaptiske område. Glutamat binder til NMDA-receptorer på de anden-ordnede neuroner, hvilket forårsager depolarisering.
Disse neuroner krydser gennem rygmarven og rejser til thalamus, hvor de synaps med tredje-ordens neuroner. Disse forbinder derefter med det limbiske system og hjernebarken. Der er også en inhiberende vej, der forhindrer smertestransmissionsoverførsel fra dorsalhornet. Anti-nociceptive neuroner stammer fra hjernestammen og rejser ned ad rygmarven, hvor de synaps med korte interneuroner i dorsalhornet ved at frigive dopamin og norepinephrin. Interneuronerne modulerer synaps mellem den første ordensneuron såvel som andenordensneuronen ved at frigive gamma-amino-smørsyre eller GABA, en inhibitorisk neurotransmitter. Følgelig er smerteafbrydelse resultatet af inhibering af synaps mellem første og anden ordens neuroner, mens smerteforøgelse kan være et resultat af undertrykkelse af hæmmende synaptiske forbindelser.
Mekanismen bag neuropatisk smerte er imidlertid ikke så klar. Flere dyreforsøg har afsløret, at mange mekanismer kan være involveret. Men man skal huske, at det, der gælder for skabninger, ikke altid gælder for mennesker. Første ordensneuroner kan øge deres affyring, hvis de er delvist beskadigede og øger mængden af natriumkanaler. Ektopiske udledninger er en konsekvens af forøget depolarisering på visse steder i fiberen, hvilket resulterer i spontan smerte og bevægelsesrelateret smerte. Inhiberende kredsløb kan blive formindsket i niveauet af dorsal horn eller hjerne stamceller, såvel som begge dele, så smerteimpulser kan rejse ubestridt.
Derudover kan der være ændringer i den centrale behandling af smerte, når det på grund af kronisk smerte og brugen af nogle lægemidler og / eller medicin, kan anden- og tredjeordens neuroner skabe en "hukommelse" af smerte og blive sensibiliseret. Der er så øget følsomhed for spinale neuroner og reducerede aktiveringstærskler. En anden teori demonstrerer begrebet sympatisk vedligeholdt neuropatisk smerte. Dette begreb blev påvist ved analgesi efter sympathektomi fra dyr og mennesker. En blanding af mekanik kan imidlertid involveres i mange kroniske neuropatiske eller blandede somatiske og neuropatiske smerteforhold. Blandt disse udfordringer i smertefeltet, og meget mere, som det vedrører neuropatisk smerte, er evnen til at kontrollere det. Der er en dobbelt komponent til dette: først vurderer kvalitet, intensitet og fremskridt; og for det andet korrekt diagnosticering af neuropatisk smerte.
Der er dog nogle diagnostiske værktøjer, som kan hjælpe klinikere med at evaluere neuropatisk smerte. Til at begynde med kan nerveledningsstudier og sensoriske fremkaldte potentialer identificere og kvantificere omfanget af skade på sensoriske, men ikke nociceptive, veje ved at overvåge neurofysiologiske reaktioner på elektriske stimuli. Derudover skrider kvantitativ sensorisk test opfattelse i reaktion på eksterne stimuli af varierende intensiteter ved at anvende stimulering til huden. Mekanisk følsomhed over for taktile stimuli måles med specialværktøjer, som von Frey-hår, pinprick med sammenlåsende nåle, såvel som vibrationsfølsomhed sammen med vibrametre og termisk smerte med termoder.
Det er også yderst vigtigt at udføre en omfattende neurologisk evaluering for at identificere motoriske, sensoriske og autonome dysfunktioner. I sidste ende er der talrige spørgeskemaer, der bruges til at skelne mellem neuropatisk smerte i nociceptiv smerte. Nogle af dem omfatter kun interviewforespørgsler (fx Neuropathic Questionnaire og ID Pain), mens andre indeholder både interviewspørgsmål og fysiske tests (fx Leeds vurdering af neuropatiske symptomer og tegn skala) og det nøjagtige nye værktøj, den standardiserede evaluering af Smerte, som kombinerer seks interviewspørgsmål og ti fysiologiske evalueringer.
Behandlingsmodaliteter for neuropatisk smerte
Farmakologiske ordninger sigter mod mekanismerne for neuropatisk smerte. Imidlertid leverer både farmakologiske og ikke-farmakologiske behandlinger fuldstændig eller delvis lindring hos næsten halvdelen af patienterne. Mange bevisbaserede udtalelser fortæller at anvende blandinger af stoffer og / eller medicin til at fungere for så mange mekanismer som muligt. Størstedelen af undersøgelserne har for det meste undersøget postherpetisk neuralgi og smertefulde diabetiske neuropatier, men resultaterne kan ikke finde anvendelse på alle neuropatiske smerteforhold.
Antidepressiva
Antidepressiva hæver synaptiske serotonin- og norepinephrinniveauer, hvorved effekten af det nedadgående analgetiske system associeres med neuropatisk smerte øges. De har været grundlæggeren af neuropatisk smertebehandling. Analgetiske virkninger kan skyldes norge-adrenalin og dopamin genoptagelsesblokade, som formodentlig forbedrer nedadgående inhibering, NMDA-receptorantagonisme og natriumkanalblokade. Tricykliske antidepressiva, såsom TCA'er; fx amitriptylin, imipramin, nortriptylin og doxepin, er stærke mod kontinuerlig smertestillende smerte sammen med spontan smerte.
Tricykliske antidepressiva har vist sig at være signifikant mere effektive til neuropatisk smerte end de specifikke serotoninreoptagelseshæmmere eller SSRI'er, såsom fluoxetin, paroxetin, sertralin og citalopram. Årsagen kan være, at de hæmmer genoptagelse af serotonin og nor-epinephrin, mens SSRI'er kun hæmmer serotonin genoptagelse. Tricykliske antidepressiva kan have ubehagelige bivirkninger, herunder kvalme, forvirring, hjerteledningsblokke, takykardi og ventrikulære arytmier. De kan også forårsage vægtøgning, reduceret anfaldstærskel og ortostatisk hypotension. Tricykliske midler skal anvendes med omhu hos ældre, som er særligt sårbare for deres akutte bivirkninger. Lægemiddelkoncentrationen i blodet skal overvåges for at undgå toksicitet hos patienter, der er langsomme medicinske metaboliseringsmidler.
Serotonin-norepinephrin reuptake hæmmere, eller SNRI'er, er en ny klasse af antidepressiva. Ligesom TCA'er synes de at være mere effektive end SSRI'er til behandling af neuropatisk smerte, fordi de også hæmmer genoptagelse af både nor-epinephrin og dopamin. Venlafaxin er lige så effektiv mod svækkende polyneuropatier, som smertefuld diabetisk neuropati, som imipramin, under henvisning til TCA, og de to er signifikant større end placebo. Ligesom TCA'erne synes SNRI'erne at give fordele uafhængige af deres antidepressive virkninger. Bivirkninger omfatter sedation, forvirring, hypertension og tilbagetrækning syndrom.
Antiepileptiske lægemidler
Antiepileptiske lægemidler kan anvendes som førstlinjebehandling specielt til visse typer neuropatisk smerte. De virker ved at modulere spændingsgatede calcium- og natriumkanaler ved at forbedre de hæmmende virkninger af GABA og ved at hæmme excitatorisk glutaminerg transmission. Antiepileptiske lægemidler er ikke påvist at være effektive til akut smerte. I kronisk smerte tilfælde synes antiepileptika kun at være effektive i trigeminal neuralgi. Carbamazepin anvendes rutinemæssigt til denne tilstand. Gabapentin, som virker ved at hæmme calciumkanalfunktionen gennem agonisthandlinger i alfa-2 delta-underenheden af calciumkanalen, er også kendt for at være effektiv til neuropatisk smerte. Gabapentin virker imidlertid centralt, og det kan forårsage træthed, forvirring og somnolens.
Ikke-opioidanalgetik
Der mangler stærke data, der understøtter brug af ikke-steroide anti-inflammatoriske lægemidler eller NSAID'er i lindring af neuropatisk smerte. Dette kan skyldes manglen på en inflammatorisk komponent til lindring af smerte. Men de er blevet udnyttet ombytteligt med opioider som adjuvanser til behandling af kræftsmerter. Der er imidlertid rapporteret komplikationer, især hos alvorligt svækkede patienter.
Opioidanalgetik
Opioid analgetika er genstand for stor debat om at lindre neuropatisk smerte. De virker ved at hæmme centrale stigende smerteimpulser. Traditionelt er neuropatisk smerte tidligere blevet observeret at være opioidresistent, hvor opioider er mere egnede metoder til koronar og somatiske nociceptive typer af smerter. Mange læger forhindrer brug af opioider til at behandle neuropatisk smerte, hovedsagelig på grund af bekymringer over stofmisbrug, afhængighed og lovgivningsmæssige problemer. Men der er mange forsøg, der har fundet opioid analgetika til at lykkes. Oxycodon var bedre end placebo til lindring af smerte, allodyni, forbedring af søvn og handicap. Opioider med kontrolleret frigivelse anbefales ifølge planlagt til patienter med konstant smerte at tilskynde konstante niveauer af analgesi, forhindre udsving i blodglukose og forhindre bivirkninger forbundet med højere dosering. Mest almindeligt anvendes orale præparater på grund af deres større brugervenlighed og omkostningseffektivitet. Transdermal, parenteral og rektal præparater anvendes generelt til patienter, som ikke kan tolerere orale lægemidler.
Lokalbedøvelse
Nærliggende anæstetika er tiltalende, fordi de takket være deres regionale indsats har minimal bivirkninger. De virker ved at stabilisere natriumkanaler på axonerne af perifere førsteordensneuroner. De fungerer bedst, hvis der kun er delvist nerveskade, og overskydende natriumkanaler har samlet sig. Aktuelt lidokain er den bedst studerede repræsentant for kurset for neuropatisk smerte. Specifikt har brugen af dette 5-procents lidokainplaster til postherpetisk neuralgi forårsaget sin godkendelse fra FDA. Patchen ser ud til at virke bedst, når der er beskadiget, men opretholdt, nociceptorfunktionen fra det perifere nervesystem fra det involverede dermatom, der demonstrerer som allodyni. Det skal indstilles direkte på det symptomatiske område i 12 timer og elimineres i yderligere 12 timer og kan bruges i mange år på denne måde. Udover lokale hudreaktioner tolereres det ofte godt af mange patienter med neuropatisk smerte.
Diverse stoffer
Clonidin, en alfa-2-agonist, viste sig at være effektiv i en delmængde af patienter med diabetisk perifer neuropati. Cannabinoider har vist sig at spille en rolle i eksperimentel smertemodulation i dyremodeller, og evidens for, at effekten akkumuleres. CB2-selektive agonister undertrykker hyperalgesi og allodyni og normaliserer nociceptive tærskler uden at fremkalde analgesi.
Interventionssmerter
Invasive behandlinger kan overvejes hos patienter, der har intractable neuropatisk smerte. Disse behandlinger omfatter epidural eller perineurale injektioner af lokalbedøvende midler eller kortikosteroider, implantation af epidural og intrathekale lægemiddelleveringsmetoder og indsættelse af rygmarvstimulatorer. Disse tilgange er forbeholdt patienter med utilsigtet kronisk neuropatisk smerte, som har mislykket konservativ medicinsk ledelse og også har oplevet grundig psykologisk evaluering. I en undersøgelse af Kim et al. Blev det vist, at en rygmarvsstimulator var effektiv til behandling af neuropatisk smerte af nerverrotets oprindelse.
Dr. Alex Jimenez's Insight
Med neuropatisk smerte opstår kroniske smertesymptomer på grund af, at nervefibrene selv er beskadiget, dysfunktionelle eller skadede, generelt ledsaget af vævsskade eller skade. Som følge heraf kan disse nervefibre begynde at sende ukorrekte smertsignaler til andre områder af kroppen. Virkningerne af neuropatisk smerte forårsaget af nervefiberskader omfatter ændringer i nervefunktion både på skadestedet og i områder omkring skaden. Forståelse af patofysiologi af neuropatisk smerte har været et mål for mange sundhedspersonale med henblik på effektivt at bestemme den bedste behandlingsmetode for at hjælpe med at håndtere og forbedre sine symptomer. Fra brug af stoffer og / eller medicin, til kiropraktisk pleje, motion, fysisk aktivitet og ernæring, kan en række behandlingsmetoder bruges til at lette neuropatisk smerte for hver enkelt persons behov.
Yderligere interventioner for neuropatisk smerte
Masser af patienter med neuropatisk smerte forfølge komplementære og alternative behandlingsmuligheder til behandling af neuropatisk smerte. Andre velkendte regimer, der anvendes til behandling af neuropatisk smerte, omfatter akupunktur, perkutan elektrisk nervestimulering, transkutan elektrisk nervestimulering, kognitiv adfærdsmæssig behandling, graderet motorbilleder og understøttende behandling og motion. Blandt disse er kiropraktisk pleje imidlertid en velkendt alternativ behandlingsmetode, der almindeligvis anvendes til at hjælpe med at behandle neuropatisk smerte. Kiropraktisk pleje, sammen med fysioterapi, motion, ernæring og livsstilsændringer kan i sidste ende tilbyde lindring for neuropatiske smerte symptomer.
Kiropraktik Care
Det er kendt, at en omfattende managementapplikation er afgørende for at bekæmpe virkningerne af neuropatisk smerte. På denne måde er kiropraktikpleje et holistisk behandlingsprogram, der kan være effektivt til forebyggelse af sundhedsmæssige problemer forbundet med nerveskader. Kiropraktisk pleje yder hjælp til patienter med mange forskellige tilstande, herunder dem med neuropatisk smerte. Lidelser af neuropatisk smerte udnytter ofte ikke-steroide antiinflammatoriske lægemidler eller NSAID'er, såsom ibuprofen eller stærke receptpligtige smertestillende midler for at lette neuropatisk smerte. Disse kan give en midlertidig løsning, men har brug for konstant brug til at håndtere smerten. Dette bidrager altid til skadelige bivirkninger og i ekstreme situationer, receptpligtig stofafhængighed.
Kiropraktisk pleje kan hjælpe med at forbedre symptomer på neuropatisk smerte og forbedre stabiliteten uden disse ulemper. En tilgang som kiropraktik pleje tilbyder et individualiseret program designet til at identificere årsagen til problemet. Gennem brugen af rygtilpasninger og manuelle manipulationer kan en kiropraktor forsigtigt korrigere eventuelle spinalforskydninger eller subluxationer, der findes langs rygsøjlens længde, hvilket kunne nedsætte konsekvenserne af nervebrydning via ombygningen af rygraden. Gendannelse af spinal integritet er afgørende for at holde et velfungerende centralnervesystem.
En kiropraktor kan også være en langsigtet behandling mod at forbedre dit generelle velbefindende. Udover spinaljusteringer og manuelle manipulationer kan en kiropraktor tilbyde næringsrådgivning, som foreskriver en kost rig på antioxidanter, eller de kan designe en fysioterapi eller træningsprogram til bekæmpelse af nervesmerter. En langsigtet tilstand kræver et langsigtet middel, og i den henseende kan en sundhedspersonale, der specialiserer sig i skader og / eller tilstande, der påvirker muskel- og nervesystemet, som en læge i kiropraktik eller kiropraktor, være uvurderlig, da de arbejder at måle gunstig forandring over tid.
Fysioterapi, motion og bevægelsesrepræsentationsteknikker har vist sig at være til gavn for neuropatisk smertebehandling. Kiropraktisk pleje tilbyder også andre behandlingsformer, som kan være nyttige til styring eller forbedring af neuropatisk smerte. Lavtliggende laserterapi, eller LLLT, har for eksempel opnået en stor fremtrædende karakter som en behandling for neuropatisk smerte. Ifølge en lang række forskningsundersøgelser blev det konkluderet, at LLLT havde positive virkninger på bekæmpelsen af analgesi til neuropatisk smerte, men der kræves yderligere undersøgelser for at definere behandlingsprotokoller, der opsummerer virkningerne af lavniveau laserterapi ved neuropatiske smertebehandlinger.
Kiropraktisk pleje omfatter også ernæringsmæssige råd, som kan hjælpe med at kontrollere symptomer forbundet med diabetisk neuropati. I en undersøgelse viste en lavfedt plantebaseret kost at forbedre glykæmisk kontrol hos patienter med type 2 diabetes. Efter ca. 20-uger i pilotundersøgelsen rapporterede de involverede personer ændringer i deres kropsvægt, og elektrokemisk hudkonduktans i foden blev rapporteret at være forbedret med interventionen. Forskningsundersøgelsen foreslog en potentiel værdi i den fedtfattige plantebaserede diætintervention for diabetisk neuropati. Desuden viste kliniske undersøgelser, at den orale påføring af magnesium L-threonat er i stand til at forebygge såvel som genoprette hukommelsesunderskud i forbindelse med neuropatisk smerte.
Kiropraktisk pleje kan også tilbyde yderligere behandlingsstrategier for at fremme nerveregenerering. Som eksempel er en forbedring af regenerering af axoner blevet foreslået for at bidrage til at forbedre funktionelt opsving efter perifer nerveskader. Elektrisk stimulering sammen med motion eller fysiske aktiviteter viste sig at fremme nerveregenerering efter forsinket nervereparation hos mennesker og rotter, ifølge nyere undersøgelser. Både elektrisk stimulering og motion var i sidste ende bestemt til at være lovende eksperimentelle behandlinger for perifere nerveskade, der synes at være klar til at blive overført til klinisk brug. Yderligere undersøgelser kan være nødvendige for fuldt ud at bestemme virkningerne af disse hos patienter med neuropatisk smerte.
Konklusion
Neuropatisk smerte er en mangfoldig enhed uden særlige retningslinjer at tage sig af. Det styres bedst ved hjælp af en tværfaglig tilgang. Smertebehandling kræver løbende evaluering, patientuddannelse, hvilket sikrer patientopfølgning og beroligelse. Neuropatisk smerte er en kronisk tilstand, der gør muligheden for den bedste behandling udfordrende. Individualisering af behandling indebærer overvejelse af indvirkningen af smerten på individets trivsel, depression og handicap sammen med efteruddannelse og evaluering. Neuropatiske smerteundersøgelser, både på molekylært niveau og i dyremodeller, er relativt nye, men meget lovende. Der forventes mange forbedringer inden for de grundlæggende og kliniske felter af neuropatisk smerte og åbner derfor døråbningerne til forbedrede eller nye behandlingsmetoder for denne deaktiverende tilstand. Omfanget af vores information er begrænset til kiropraktik såvel som til rygskader og tilstande. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emner: Rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og savnede dage på arbejdspladsen over hele verden. Faktisk er rygsmerter blevet tilskrevet som den næst mest almindelige årsag til doktorkontorbesøg, der kun overstiger luftvejsinfektioner. Ca. 80 procent af befolkningen vil opleve en form for rygsmerter mindst én gang i hele deres liv. Ryggraden er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette skader og / eller forværrede forhold, som f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
Smerte Angst Depression�Alle har oplevet smerte, men der er dem med depression, angst eller begge dele. Kombiner dette med smerte, og det kan blive ret intenst og svært at behandle. Mennesker, der lider af depression, angst eller begge dele, har en tendens til at opleve svær og langvarig smerte mere end andre mennesker.
Vejen angst, depression og smerter overlapper hinanden ses ved kroniske og ved nogle invaliderende smertesyndromer, dvs. lænderygsmerter, hovedpine, nervesmerter og fibromyalgi. Psykiatriske lidelser bidrager til smerteintensiteten og øger også risikoen for invaliditet.
Depression:A (major depressiv lidelse eller klinisk depression) er en almindelig, men alvorlig stemningsforstyrrelse. Det forårsager alvorlige symptomer, der påvirker, hvordan en person føler, tænker og hvordan den håndterer daglige aktiviteter, dvs. at sove, spise og arbejde. For at blive diagnosticeret med depression skal symptomerne være til stede i mindst to uger.
Vedvarende trist, ængstelig eller 'let' humør.
Følelser af håbløshed, pessimistisk.
Irritabilitet.
Følelser af skyld, værdiløshed eller hjælpeløshed.
Tab af interesse eller fornøjelse i aktiviteter.
Mindsket energi eller træthed.
Flytter eller taler langsomt.
Føler mig rastløs og har problemer med at sidde stille.
Vanskeligheder koncentrere, huske eller træffe beslutninger.
Sværhedsvanskeligheder, opvågnen tidligt om morgenen og sovende.
Appetit og vægtændringer.
Tanker om død eller selvmord & eller selvmordsforsøg.
Hæv eller smerter, hovedpine, kramper eller fordøjelsesproblemer uden en klar fysisk årsag og / eller som ikke lette behandlingen.
Ikke alle, der er deprimerede, oplever hvert eneste symptom. Nogle oplever kun få symptomer, mens andre kan opleve flere. Adskillige vedvarende symptomer ud over dårligt humør er�påkrævet�til en diagnose af svær depression. Sværhedsgraden og hyppigheden af symptomer sammen med varigheden vil variere afhængigt af den enkelte og deres særlige sygdom. Symptomerne kan også variere afhængigt af sygdommens stadium.
SMERTE ANGST DEPRESSION
mål:
Hvad er forholdet?
Hvad er neurofysiologien bag det?
Hvad er de centrale konsekvenser?
Hjerneændringer i smerte
Figur 1 Hjernebaner, regioner og netværk involveret i akutte og kroniske smerter
Davis, KD et al. (2017) Hjernebilleddiagnostiske tests for kroniske smerter: medicinske, juridiske og etiske spørgsmål og anbefalinger Nat. Rev. Neurol. doi:10.1038/nrneurol.2017.122
SMERTE, ANGST OG DEPRESSION
konklusion:
Smerter, især kroniske, er forbundet med depression og angst
De fysiologiske mekanismer, der fører til angst og depression, kan være af multifaktoriel natur
Smerter forårsager ændringer i hjernen struktur og funktion
Denne ændring i struktur og funktion kan ændre hjernens evne til at modulere smerte samt kontrollere humør.
Når det sensoriske system er påvirket af skade eller sygdom, kan nerverne i det pågældende system ikke fungere korrekt for at overføre følelser i hjernen. Dette bidrager ofte til følelse af følelsesløshed eller mangel på sensation. Men i visse tilfælde, når dette system er beskadiget, kan folk opleve smerter i det berørte område.
neuropatisk smerte begynder ikke brat eller løser hurtigt; Det er en kronisk smerte tilstand, der fører til vedvarende smerte symptomer. For de fleste individer kan intensiteten af deres symptomer vokse og aftage hele dagen. Selvom neuropatisk smerte skal relateres til perifere nervehelseproblemer, som neuropati forårsaget af diabetes eller spinalstenose, kan skader på hjernen eller rygmarven også føre til kronisk neuropatisk smerte. Neuropatisk smerte kaldes også nervesmerter.
Neuropatisk smerte kan modvirke nociceptiv smerte. Neuropatisk smerte udvikler sig ikke til nogen specifik omstændighed eller udefrakommende stimulus, men symptomerne opstår simpelthen fordi nervesystemet måske ikke virker i overensstemmelse hermed. Faktisk kan enkeltpersoner også opleve neuropatisk smerte, selvom den smertefulde eller skadede kropsdel ikke er der faktisk. Denne tilstand kaldes phantom limb smerte, som kan forekomme hos mennesker, efter at de har haft en amputation.
Nociceptiv smerte er generelt akut og udvikler sig som reaktion på en bestemt omstændighed, som når en person oplever en pludselig skade, som at hamre en finger med en hammer eller stubbende tå, når han går barfodet. Desuden har nociceptive smerter tendens til at gå væk, når det berørte sted helbreder. Kroppen indeholder specialiserede nerveceller, kendt som nociceptorer, som opdager skadelige stimuli, som kan skade kroppen, såsom ekstrem varme eller kulde, tryk, klemning og eksponering for kemikalier. Disse advarselssignaler sendes derefter langs nervesystemet til hjernen, hvilket resulterer i nociceptiv smerte.
Hvad er risikofaktorer for neuropatisk smerte?
Alt, der bidrager til manglende funktion inden for det sensoriske nervesystem, kan føre til neuropatisk smerte. Som sådan kan nervesygdomme fra karpaltunnelsyndrom eller lignende tilstande i sidste ende udløse neuropatisk smerte. Traume, der resulterer i nerveskade, kan føre til neuropatisk smerte. Andre tilstande, som kan foreskrive personer til udvikling af neuropatisk smerte, omfatter: diabetes, vitaminmangel, kræft, hiv, slagtilfælde, multipel sklerose, helvedesild og endda nogle kræftbehandlingsformer.
Hvad er årsagerne til neuropatisk smerte?
Der er mange årsager, hvorfra personer kan udvikle neuropatisk smerte. Men på et cellulært niveau er en forklaring en forøget frigivelse af bestemte receptorer, der indikerer smerte, sammen med en nedsat evne af nerverne til at modulere disse signaler fører til følelsen af smerte, der stammer fra den berørte region. Desuden er rygsøjlen i rygmarven omorganiseret med tilsvarende ændringer i hormoner og tab af normalt fungerende mobile legemer. Disse ændringer resulterer i opfattelsen af smerte i mangel af ekstern stimulering. I hjernen kan evnen til at blokere smerte blive påvirket efter en skade, såsom slagtilfælde eller traume fra en skade. Som tiden går, sker der yderligere celleskader, og følelsen af smerte fortsætter. Neuropatisk smerte er også relateret til diabetes, kronisk alkoholindtagelse, visse kræftformer, vitamin B-mangel, sygdomme, andre nerverelaterede sygdomme, toksiner og specifikke lægemidler.
Hvad er symptomerne på neuropatisk smerte?
I modsætning til andre neurologiske tilstande kan identifikation af neuropatisk smerte være udfordrende. Imidlertid kan flere, hvis nogen objektive signaler er til stede. Sundhedspersonale skal dechifrere og oversætte et udvalg af ord, som patienter bruger til at beskrive deres smerte. Patienterne kan beskrive deres symptomer som skarpe, kedelige, varme, kolde, følsomme, kløende, dybe, stikkende, brændende, blandt en række andre beskrivende termer. Derudover kan nogle patienter opleve smerte ved let tryk eller tryk.
I et forsøg på at hjælpe med at identificere, hvor meget smerte patienter kan gennemgå, anvendes forskellige skalaer ofte. Patienterne bliver bedt om at bedømme deres smerte i henhold til en visuel skala eller numerisk graf. Der findes mange eksempler på smerteskalaer, som den der er vist nedenfor. Ofte kan billeder af ansigter, der viser forskellige niveauer af smerte, være nyttige, når personer har en vanskelig tid, der beskriver mængden af smerte, de oplever.
Kronisk smerte og mental sundhed
For mange kan virkningen af kronisk smerte ikke være begrænset til smerten; det kan også påvirke deres mentale tilstand negativt. Nye forskningsundersøgelser foretaget af forskere ved Northwestern University i Chicago kan forklare, hvorfor personer, der har kronisk smerte, også lider af tilsyneladende ikke-relaterede sundhedsspørgsmål, såsom depression, stress, søvnløshed og problemer med at koncentrere sig.
Evalueringen viste, at personer med kronisk smerte viser forskellige regioner i hjernen, som altid er aktive, især området der er forbundet med humør og opmærksomhed. Denne kontinuerlige handling genskaber nerveforbindelser fra hjernen og efterlader kroniske smertepasienter med større risiko for psykiske problemer. Forskere foreslog at få smertsignaler hele tiden kunne resultere i mental genskabelse, der har negativ indflydelse på sindet. Rewiring tvinger deres hjerner til at bruge mentale ressourcer forskelligt til at håndtere hverdagens opgaver, fra matematik, til at mindes en indkøbsliste, for at føle sig lykkelig.
Smertehjerneforbindelsen har været velindspillet, i det mindste anekdotisk, og mange sundhedspersonale siger, at de har set førstehånds, hvordan patientens mentale tilstand kan gå ned ad bakke, når de udviser kronisk smerte. Misforståelser om smerte-hjerneforbindelsen kan være opstået ud fra manglende bevis for, at smerte har en målbar, vedvarende indflydelse på hjernen. Forskere forventer, at med yderligere undersøgelse af mekanismerne for, hvordan kronisk smerte gør folk mere modtagelige for humørsygdomme, vil folk være i stand til bedre at klare deres generelle trivsel.
Kultur og kronisk smerte
Mange ting bidrager til den måde, vi oplever og udtrykker smerte på, men det er også for nylig blevet foreslået af forskere, at kultur direkte vedrører udtryk for smerte. Vores opdragelse og samfundsmæssige værdier påvirker, hvordan vi udtrykker smerte og også dets egen natur, intensitet og længde. Disse variabler er imidlertid ikke så indlysende som socio-psykologiske værdier, såsom alder og køn.
Forskning siger, at kronisk smerte er en multifacetteret proces og det samtidige samspil mellem patofysiologi, kognitive, affektive, adfærdsmæssige og sociokulturelle faktorer summerer til det, der kaldes den kroniske smerteoplevelse. Det fremgår, at kronisk smerte opleves forskelligt blandt patienter af forskellige kulturer og etniske grupper.
Nogle kulturer tilskynder til udtryk for smerte, især i det sydlige Middelhav og Mellemøsten. Andre personer undertrykker det, som i de mange lektier til vores børn om at opføre sig modigt og ikke græde. Smerter er anerkendt som en del af den menneskelige oplevelse. Vi er tilbøjelige til at antage, at kommunikation om smerte nøje vil krydse kulturelle grænser. Men mennesker i smerte er underlagt de manerer, deres civilisationer har uddannet dem til at opleve og udtrykke smerte.
Begge personer i smerte og sundhedspersonale oplever vanskeligheder med at kommunikere smerte over de etniske grænser. I en sag som smerte, hvor effektiv kommunikation kan have vidtrækkende konsekvenser for lægebehandling, livskvalitet og potentiel overlevelse, bliver kulturens rolle i smertekommunikation fortsat underevalueret. Vedvarende smerte er et flerdimensionalt, sammensat møde dannet ved sammenvævning og medvirkning af biologiske og psykosociale faktorer. At kende kulminationen af disse faktorer er afgørende for at forstå forskellene i dens manifestation og ledelse.
Hvordan diagnosticeres neuropatisk smerte?
Diagnosen af neuropatisk smerte er afhængig af yderligere vurdering af en persons historie. Hvis der er mistanke om underliggende nerveskader, kan analyse af nerverne sammen med testning være berettiget. Det mest almindelige middel til at vurdere, om en nerve er skadet, bruger elektrodiagnostisk medicin. Denne medicinske subspecialty anvender teknikker for nerveledningsstudier med elektromyelografi (NCS / EMG). Klinisk evaluering kan vise tegn på tab af arbejde og kan omfatte evaluering af lysfølsomhed, evnen til at skelne skarpt ud af kedelig smerte og evnen til at skelne mellem temperatur og evaluering af vibrationer.
Efter en grundig klinisk undersøgelse er afsluttet, kan den elektrodiagnostiske analyse planlægges. Disse undersøgelser udføres af en specialuddannet neurolog og fysiater. Hvis der er mistanke om neuropati, bør en jagt på reversible årsager udføres. Dette kan omfatte blodfunktion for mangel på vitamin eller skjoldbruskkirtlen og billeddannelsesundersøgelser for at udelukke en strukturel læsion, som påvirker rygmarven. Afhængigt af resultaterne af denne test kan der være et middel til at reducere intensiteten af neuropati og muligvis reducere smerten, som en patient undergår.
Desværre kan selv i mange tilfælde ikke en god kontrol med den underliggende årsag til neuropati reversere den neuropatiske smerte. Dette ses almindeligvis hos patienter med diabetisk neuropati. I sjældne tilfælde kan der være tegn på ændringer i hud- og hårvækstmønsteret i en berørt region. Disse ændringer kan være forbundet med ændringer i sved. Hvis de er til stede, kan disse ændringer hjælpe med at identificere den sandsynlige forekomst af neuropatisk smerte relateret til en tilstand kendt som komplekst regionalt smertesyndrom.
Dr. Alex Jimenez's Insight
Neuropatisk smerte er en kronisk smerte tilstand, som generelt er forbundet med direkte skade eller skade på nervesystemet eller nerverne. Denne type smerte er forskellig fra nociceptiv smerte, eller den typiske følelse af smerte. Nociceptiv smerte er en akut eller pludselig følelse af smerte, der forårsager, at nervesystemet sender signaler af smerte umiddelbart efter traumet. Med neuropatisk smerte kan patienterne dog opleve skud, brændende smerter uden direkte skader eller skade. At forstå de mulige årsager til patientens neuropatiske smerter i forhold til enhver anden form for smerte, kan hjælpe sundhedspersonale med at finde bedre måder at behandle kroniske smerteforhold.
Hvad er behandlingen for neuropatisk smerte?
Forskellige lægemidler anvendes i et forsøg på at behandle neuropatisk smerte. Størstedelen af disse lægemidler anvendes off-label, hvilket betyder, at lægemidlet blev godkendt af FDA til behandling af forskellige tilstande og blev derefter anerkendt som værende fordelagtig til behandling af neuropatisk smerte. Tricykliske antidepressiva, såsom amitriptylin, nortriptylin og desipramin, er blevet ordineret til behandling af neuropatisk smerte i flere år.
Nogle personer finder, at disse kan være meget effektive til at give dem lindring. Andre former for antidepressiva har vist sig at give en vis lettelse. Selektive serotoninoptagelseshæmmere eller SSRI'er, såsom paroxetin og citalopram, og andre antidepressiva, såsom venlafaxin og bupropion, er blevet anvendt hos visse patienter. En anden hyppig behandling af neuropatisk smerte inkorporerer antiseptiske lægemidler, herunder carbamazepin, phenytoin, gabapentin, lamotrigin og andre.
I akutte tilfælde af smertefuld neuropati, der ikke reagerer på førstegangsmæglere, kan stoffer, der typisk anvendes til behandling af hjertearytmi, være til nytte; Men disse kan føre til betydelige bivirkninger og skal ofte overvåges nøje. Medicin, der påføres direkte på huden, kan give beskedne fordele for nogle patienter. De almindeligt anvendte former omfatter lidokain (i patch eller geltype) eller capsaicin.
Behandling af neuropatisk smerte er afhængig af den underliggende årsag. Hvis årsagen er reversibel, kan perifere nerver regenerere og smerten vil aftage Ikke desto mindre kan denne reduktion i smerte tage flere måneder til år. Adskillige andre alternative behandlingsmuligheder, herunder kiropraktikpleje og fysioterapi, kan også bruges til at hjælpe med at lindre spændinger og stress langs nerverne og i sidste ende bidrage til at forbedre smertefulde symptomer.
Hvad er prognosen for neuropatisk smerte?
Mange personer med neuropatisk smerte er i stand til at få en vis grad af hjælp, selv når deres smerte vedvarer. Selv om neuropatisk smerte ikke er farlig for en patient, kan tilstedeværelsen af kronisk smerte påvirke livskvaliteten negativt. Patienter med kronisk nervepine kan lide under søvnmangel eller humørsygdomme, herunder depression, angst og stress, som tidligere nævnt. På grund af den iboende alopeci og mangel på sensorisk tilbagekobling er patienter i fare for at udvikle skade eller infektion eller ubevidst forårsager en eskalering af en nuværende skade. Derfor er det vigtigt at søge øjeblikkelig lægehjælp og følge specifikke retningslinjer udarbejdet af en sundhedspersonel for sikkerhed og forsigtighed.
Kan neuropatisk smerte forhindres?
Den bedste måde at forhindre neuropatisk smerte på er at undgå udvikling eller progression af neuropati. Overvågning og ændring af livsstilsindstillinger, herunder begrænsning af brugen af alkohol og tobak holde en sund vægt for at mindske risikoen for diabetes, degenerativ ledsygdom eller slagtilfælde; og at have en god ergonomisk form på arbejdspladsen eller når man praktiserer hobbyer for at mindske risikoen for gentagne stressskader er strategier for at mindske sandsynligheden for at udvikle neuropati og potentiel neuropatisk smerte. Sørg for at søge øjeblikkelig lægehjælp i tilfælde af symptomer forbundet med neuropatisk smerte for at fortsætte med den mest hensigtsmæssige behandlingsmetode. Omfanget af vores information er begrænset til kiropraktik såvel som til rygmarvsskader og tilstande. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emner: Rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og savnede dage på arbejdspladsen over hele verden. Faktisk er rygsmerter blevet tilskrevet som den næst mest almindelige årsag til doktorkontorbesøg, der kun overstiger luftvejsinfektioner. Ca. 80 procent af befolkningen vil opleve en form for rygsmerter mindst én gang i hele deres liv. Ryggraden er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette skader og / eller forværrede forhold, som f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
Hvis det sensoriske system påvirkes af skade eller sygdom, kan nerverne i systemet ikke fungere i transmissionen af sensation til hjernen. Dette kan føre til en følelse af følelsesløshed eller manglende fornemmelse. I nogle tilfælde når det sensoriske system er skadet, kan enkeltpersoner opleve smerter i det berørte område. Neuropatisk smerte starter ikke hurtigt eller slutter hurtigt. Det er en kronisk tilstand, der fører til symptomer på vedvarende smerter. For mange kan intensiteten af symptomerne komme og gå igennem en dag. Neuropatisk smerte menes at være forbundet med perifere nerveproblemer, dvs. neuropati forårsaget af diabetes, spinal stenose, skade på hjernen eller rygmarven kan også føre til kronisk neuropatisk smerte.
NEUROPATISKE PAIN
mål:
Hvad er det?
Hvad er patofysiologien bag den?
Hvad er årsagerne til
Hvad er nogle af veje
Hvordan kan vi løse det?
NEUROPATISKE PAIN
Smerte initieret eller forårsaget af en primær læsion eller dysfunktion i det somatosensoriske nervesystem.
Neuropatisk smerte er normalt kronisk, vanskelig at behandle og ofte resistent over for standard analgetisk behandling.
PATHOGENESIS AF NEUROPATISK PAIN
PERIPHERAL MECHANISMS
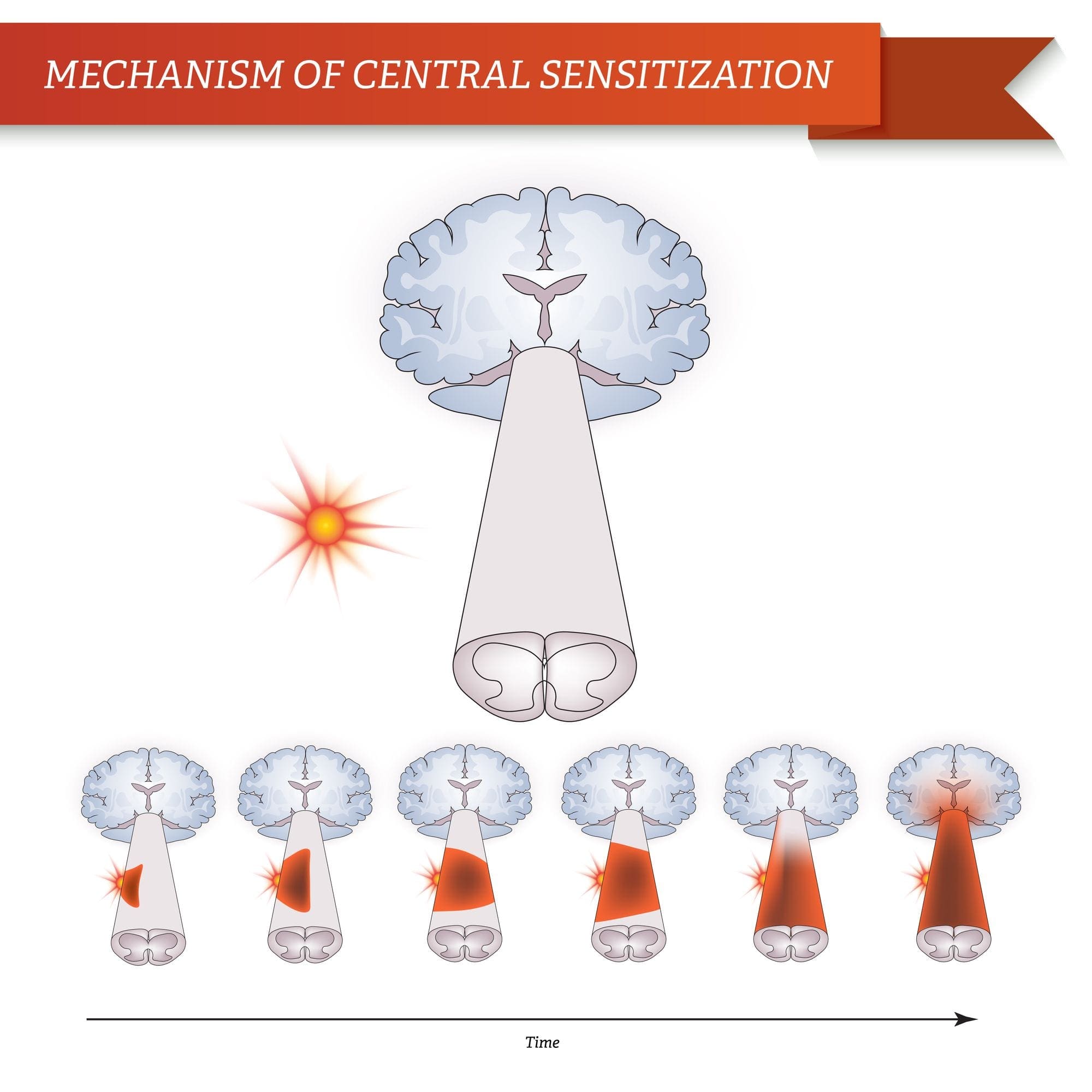
Efter en perifert nerve læsion bliver neuroner mere følsomme og udvikler unormal excitabilitet og forhøjet sensitivitet til stimulering
Dette kaldes ... Perifere sensibilisering!
CENTRALE MEKANISMER
Som en konsekvens af den igangværende spontane aktivitet, som opstår i periferien, udvikler neuroner en øget baggrundsaktivitet, forstørrede modtagelige felter og øgede reaktioner på afferente impulser, herunder normale taktile stimuli
Dette er kendt som ...Central sensibilisering!
Almindelige årsager
Lesioner eller sygdomme i det somatosensoriske nervesystem kan føre til forandret og uordnet transmission af sensoriske signaler i rygmarven og hjernen; Almindelige tilstande forbundet med neuropatisk smerte omfatter:
Postherpetic neuralgi
Trigeminal neuralgi
Smertefuld radikulopati
Diabetisk neuropati
HIV-infektion
Spedalskhed
Amputation
Perifere nerve skadesmerter
Stroke (i form af central post-stroke smerte)
PHANTOM LEMMESMÆRKE & AUSTERET VIRKELIGHED
Phantom Limb Pain og AR
NEUROGENISK INFLAMMATION
mål:
Hvad er det?
Hvad er patofysiologien bag den?
Hvad er årsagerne til
Hvordan kan vi løse det?
NEUROGENISK INFLAMMATION
Neurogen inflammation er en neuralt fremkaldt lokal inflammatorisk respons karakteriseret ved vasodilation, øget vaskulær permeabilitet, mastcelle degranulation og frigivelse af neuropeptider inklusive SP og calcitonin-genrelateret peptid (CGRP)
Det ser ud til at spille en vigtig rolle i patogenesen af talrige sygdomme, herunder migræne, psoriasis, astma, fibromyalgi, eksem, rosacea, dystoni og flere kemiske følsomheder
Almindelige årsager
Der er flere veje, hvorved neurogen inflammation kan initieres. Det er veldokumenteret med anvendelse af både dyremodeller og isolerede neuroner in vitro, at capsaicin, varme, protoner, bradykinin og tryptase er opstrøms regulatorer af den intracellulære calciumstrøm, hvilket resulterer i frigivelse af inflammatorisk neuropeptid. I modsætning hertil antages det, at prostaglandiner E2 og I2, cytokiner, interleukin-1, interleukin-6 og tumornekrosefaktoren ikke forårsager neurotransmitter frigørelse af sig selv, men snarere ophidser sensoriske neuroner og dermed sænker tærsklen for affyring og forårsager forstærket frigivelse af neuropeptider.
Mens neurogen inflammation er blevet grundigt undersøgt og veldokumenteret i perifere væv, har konceptet neurogent inflammation inden for CNS indtil videre været stort set uudforsket. I betragtning af kapaciteten til neurogen inflammation for at påvirke vaskulær permeabilitet og føre til dannelse af ødem, er det nu blevet undersøgt meget for dets potentiale til at påvirke BBB permeabilitet og vasogent ødem i hjernen og rygmarven under forskellige patologiske tilstande.
Vi blev alle undervist som børn, at der er 5 sanser: syn, smag, lyd, lugt og berøring. De første fire sanser bruger klare, adskilte organer, som f.eks. Øjne, smagsløg, ører og næse, men hvordan virker kropssansen nøjagtigt? Touch er oplevet over hele kroppen, både indenfor og udenfor. Der er ikke et særskilt organ, der er ansvarlig for at føle berøring. Der er snarere små receptorer eller nerveender rundt omkring i hele kroppen, som fornemmer at røre ved hvor det forekommer og sender signaler til hjernen med oplysninger om den type berøring der opstod. Som smagsprøve på tungen opdager smag er mekanoreceptorer kirtler i huden og på andre organer, der registrerer følelser af berøring. De er kendt som mekanoreceptorer fordi de er designet til at opdage mekaniske fornemmelser eller forskelle i tryk.
Rolle af mekanoreceptorer
En person forstår, at de har oplevet en fornemmelse, når organet der er ansvarlig for at opdage, at den specifikke forstand sender en besked til hjernen, som er det primære organ, som behandler og arrangerer al information. Meddelelser sendes fra alle områder af kroppen til hjernen gennem ledninger benævnt neuroner. Der er tusindvis af små neuroner, der forgrener sig til alle områder af menneskekroppen, og på slutningen af mange af disse neuroner er mekanoreceptorer. For at demonstrere, hvad der sker, når du rører et objekt, bruger vi et eksempel.
Forestil dig en mygeland på din arm. Stammen af dette insekt, så let, stimulerer mekanoreceptorer i det pågældende område af armen. Disse mekanoreceptorer sender en besked langs den neuron, de er forbundet med. Neuronet forbinder hele vejen til hjernen, som modtager meddelelsen om, at noget berører din krop i den nøjagtige placering af den specifikke mekanoreceptor, der sendte beskeden. Hjernen vil handle med dette råd. Måske vil det fortælle øjnene at se på den region af armen, der registrerede signaturen. Og når øjnene fortæller hjernen, at der er en myg på armen, kan hjernen fortælle hånden om hurtigt at smide det væk. Sådan virker mekanoreceptorer. Formålet med artiklen nedenfor er at demonstrere og diskutere i detaljer den funktionelle organisering og molekylære determinanter for mekanoreceptorer.
Touch Sense: Funktionel organisation og molekylære determinanter af mekanosensitive receptorer
Abstrakt
Kutane mekanoreceptorer er lokaliseret i de forskellige lag i huden, hvor de opdager en bred vifte af mekaniske stimuli, herunder lys børste, stræk, vibration og skadeligt tryk. Denne variation af stimuli matches af et mangfoldigt udvalg af specialiserede mekanoreceptorer, der reagerer på kutan deformation på en bestemt måde og relæer disse stimuli til højere hjernestrukturer. Undersøgelser på tværs af mekanoreceptorer og genetisk indgreb i sensoriske nerveender begynder at afdække touch sensationsmekanismer. Arbejdet på dette område har givet forskere en mere grundig forståelse af kredsløbsorganisationen, der ligger til grund for opfattelsen af berøring. Nye ionkanaler er opstået som kandidater til transduktionsmolekyler og egenskaber af mekanisk gatedstrømme forbedrede vores forståelse af mekanismerne til tilpasning til taktile stimuli. Denne gennemgang fremhæver de fremskridt, der er gjort med at karakterisere funktionelle egenskaber hos mekanoreceptorer i hårede og glabro hud- og ionkanaler, der registrerer mekaniske indgange og form mekanoreceptor tilpasning.
Touch er påvisning af mekanisk stimulering, der påvirker huden, herunder uskadelige og skadelige mekaniske stimuli. Det er en væsentlig fornuft for overlevelse og udvikling af pattedyr og mennesker. Kontakt med faste genstande og væsker med huden giver nødvendig information til centralnervesystemet, der muliggør efterforskning og anerkendelse af miljøet og initierer bevægelse eller planlagt håndbevægelse. Touch er også meget vigtigt for lærlingeforhold, sociale kontakter og seksualitet. Berøringsfølsomhed er den mindst sårbare fornemmelse, selv om den kan forvrænges (hyperesthesi, hypoestesi) under mange patologiske forhold. 1-3
Berøringssvar involverer en meget præcis kodning af mekanisk information. Kutane mekanoreceptorer er lokaliseret i de forskellige lag af huden, hvor de registrerer en bred vifte af mekaniske stimuli, herunder let børste, strækning, vibration, afbøjning af hår og skadeligt tryk. Denne række stimuli matches af en række forskellige specialiserede mekanoreceptorer, der reagerer på kutan deformation på en bestemt måde og videreformidler disse stimuli til højere hjernestrukturer. Somatosensoriske neuroner i huden falder i to grupper: mekanismer med lav tærskel (LTMR'er), der reagerer på godartet tryk, og højterskel mekanoreceptorer (HTMR'er), der reagerer på skadelig mekanisk stimulering. LTMR- og HTMR-cellekroppe befinder sig i dorsal rodganglier (DRG) og kraniale sensoriske ganglier (trigeminusganglier). Nervefibre associeret med LTMR'er og HTMR'er klassificeres som Aa -, Aa - eller C-fibre baseret på deres handlingspotentiale ledningshastigheder. C-fibre er umyeliniserede og har de langsomste ledningshastigheder (~ 2 m / s), mens A? og A? fibre er let og stærkt myeliniserede og udviser henholdsvis mellemliggende (~ 12 m / s) og hurtige (~ 20 m / s) ledningshastigheder. LTMR'er klassificeres også som langsomt eller hurtigt tilpasningsresponser (SA- og RA-LTMR'er) i henhold til deres tilpasningshastigheder til vedvarende mekanisk stimulus. De adskiller sig yderligere ved de kutane endeorganer, de innerverer, og deres foretrukne stimuli.
Mulighed for mekanoreceptorer til at detektere mekaniske signaler er afhængig af tilstedeværelsen af mekanotransducer ionkanaler, der hurtigt omdanner mekaniske kræfter til elektriske signaler og depolariserer det modtagelige felt. Denne lokale depolarisation, der kaldes receptorpotentiale, kan generere actionpotentialer, der formerer sig mod centralnervesystemet. Egenskaber af molekyler, der formidler mekanotransduktion og tilpasning til mekaniske kræfter forbliver uklare.
I denne anmeldelse giver vi en oversigt over pattedyrs mekanoreceptoregenskaber i uskadelig og skadelig berøring i den hårede og glat hud. Vi overvejer også den nylige viden om egenskaberne ved mekanisk styrede strømme i et forsøg på at forklare mekanismen til mekanoreceptors tilpasning. Endelig gennemgår vi de seneste fremskridt med at identificere ionkanaler og tilknyttede proteiner, der er ansvarlige for genereringen af mechano-gatede strømme.
Innocuous Touch
Hårfollikel-associerede LTMR'er
Hårsækkene repræsenterer hårsaksproducerende mini-organer, der opdager let berøring. Fibre, der er forbundet med hårsækkene, reagerer på hårbevægelse og dets retning ved at skyde tog af handlingspotentialer ved begyndelsen og fjernelsen af stimulus. De tilpasser hurtigt receptorer.
Katte og kanin. I katte- og kaninfrakke kan hårsækkene opdeles i tre hårsækketyper, Down-hår, Guard-hår og Tylotrichs. Nedhårene (underhår, uld, vellus) 4 er de mest talrige, de korteste og fineste hår i pelsen. De er bølgede, farveløse og fremkom i grupper på to til fire hår fra en almindelig åbning i huden. Vagthårene (monotrichs, overhør, tophair) 4 er let buede, enten pigmenterede eller upigmenterede og kom enkeltvis ud af munden på deres follikler. Tylotrichs er de mindst talrige, de længste og tykkeste hår.5,6 De er pigmenterede eller upigmenterede, nogle gange begge og kom enkeltvis ud af en follikel, der er omgivet af en løkke med kapillære blodkar. De sensoriske fibre, der leveres til en hårsække, er placeret under talgkirtlen og tilskrives A? eller A? -LTMR fibre.7
I umiddelbar nærhed til den nedre håraksel er lige under niveauet af talgkirtlen ringet af lancetformede pilo-Ruffini-slutninger. Disse sensoriske nerveender er anbragt i en spiralbane rundt om hårets aksel i bindevævet, der danner hårsækken. Inden for hårsækken er der også frie nerveender, hvoraf nogle danner mekanoreceptorer. Hyppigt, berør corpuscles (se glabro hud) omkring halsområdet af tylotrich follikel.
Egenskaber for myeliniserede nerveender i hår af kat og kanin er blevet udforsket intenst i perioden 1930-1970 (gennemgang i Hamann, 1995). 8 Bemærkelsesværdigt, Brown og Iggo, der studerede 772 enheder med myeliniserede afferente nervefibre i de saphenøse nerver fra kat og kanin, har klassificeret respons i tre receptortyper svarende til bevægelserne af Down-hår (type D-receptorer), Guard-hår (type G-receptorer) og Tylotrich-hår (type T-receptor) .9 Alle de afferente nervefibersvar er blevet samlet i den hurtigt tilpasningsreceptor af type I (RA I) ved modstand mod den Pacinian-receptor ved navn RA II. RA I mekanoreceptorer registrerer hastigheden af mekanisk stimulus og har skarp kant. De registrerer ikke termiske variationer. Burgess et al. beskrev også en hurtigt tilpasende feltreceptor, der reagerer optimalt på strøgning af huden eller bevægelse af flere hår, hvilket blev tilskrevet stimulering af pilo-Ruffini-ender. Intet af hårsækkets respons blev tilskrevet C-fiberaktivitet.10
Mus. I musens dorsale hårde hud er tre hovedtyper af hårsække blevet beskrevet: zigzag (omkring 72%), awl / auchene (omkring 23%) og vagt eller tylotrich (omkring 5%). 11-14 Zigzag og Awl / auchenne hårsække producerer de tyndere og kortere håraksler og er forbundet med en talgkirtlen. Guard eller tylotrich hår er den længste af hårsække typer. De er præget af en stor hårpære forbundet med to talgkirtler. Vagt og A / S hår er arrangeret i et iterativt, regelmæssigt mellemrummet mønster, mens zigzaghår tætbefolket hudområder omkring de to større hårfollikeltyper [Fig. 1 (A1, A2 og A3)].
Figur 1. Organisation og fremskrivninger af kutane mekanoreceptorer. I hårig hud opdages lyspensel og berøring hovedsageligt af innerveringen omkring hårsækkene: awl / auchenne (A1), zigzag (A2) og vagt (A3). Awl / auchene-hår er tredobbelt innerveret af C-LTMR-lancetformede ender (A4), A? -LTMR og A? hurtigt tilpasning-LTMR (A6). Zigzag-hårsækkene er de kortere håraksler og er innerveret af både C-LTMR (A4) og A? -LTMR lancetformede ender (A5). De længste vagt hårsækkene er innerveret af A? hurtigt tilpasning-LTMR langsgående lancetformede ender (A6) og er forbundet med A? langsomt tilpasning-LTMR af touch dome-ender (A7). De centrale fremspring af alle disse fibre slutter i distinkte, men delvist overlappende laminater i rygmarvets dorsale horn (C-LTMR i lamina II, Aa-LTMR i lamina III og Aa-LTMR i lamina IV og V). Fremspringene af LTMR, som innerverer de samme eller tilstødende hårsækkede stoffer, er justeret for at danne en smal søjle i rygmarvets dorsale horn (B1 i grå). Kun i hårig hud indvander en subpopulation af C-fibre frie ende epidermis og reagerer på behagelige berøring (A8). Disse C-touch-fibre reagerer ikke på skadelig berøring, og deres vejkørsel er endnu ikke kendt (B2). I glabro hud medieres uskadelig berøring af fire typer LTMR'er. Merkel-celle-neurit-komplekset er i det basale lag af epidermis (C1). Denne mekanoreceptor består af et arrangement mellem mange Merkel-celler og en forstørret nerveterminal fra en enkelt A? fiber. Merkelceller udviser fingerlignende processer, der kontakter keratinocytter (C2). Ruffini-slutningen er lokaliseret i dermis. Det er en tynd cigarformet indkapslet sensorisk slutning forbundet med A? fiber (C3). Meissner-legemet forbundet med A? nerveender og er placeret i dermale papiller. Denne indkapslede mekanoreceptor består af pakket ned understøttende celler indrettet som vandrette lameller omgivet af bindevæv (C4). Pacinian corpuscle er den dybere mekanoreceptor. En enkelt A? umyelineret nerveende slutter i midten af denne store ovoide blodlegeme lavet af koncentriske lameller. Fremskrivninger af disse A? -LTMR-fibre i rygmarven er opdelt i to grene. Den vigtigste centrale gren (B3) stiger i rygmarven i den ipsilaterale dorsale danner cuneate eller gracile hæfterne (B5) upon medulla niveau, hvor de primære afferenter gør deres første synapse (B6). De sekundære neuroner gør en sensorisk decussation (B7) til dannelse af en kanal på den mediale lemniscus, der stiger gennem hjernestammen til midterbenet, specielt i thalamus. Den sekundære branche af LTMR ophører i dorsalhornet i lamina II, IV, V og forstyrrer smertetransmissionen (B4). Kræftende berøring opdages af den frie nerve ende i epidermis af begge hårede (A9) og glabro hud (C7). Disse mekanoreceptorer er afslutningen på Aa-HTMR og C-HTMR i tæt kontakt med nærliggende keratinocytter (C6). Aa-hTMR slutter i lamellen I og V; C-HTMR afsluttes i lamina I og II (B8). På rygmarvsdorsalhornsniveau producerer primære afferenter HTMR synapser med sekundære neuroner, som krydser midterlinien og klatre op til den højere hjernestruktur i den anterolaterale fascikel (B9, B10).
For nylig brugte Ginty og samarbejdspartnere en kombination af molekylær-genetisk mærkning og somatotopiske retrograd sporingsmetoder til at visualisere organisationen af perifere og centrale aksonale ender af LTMR'erne hos mus.15 Deres fund understøtter en model, hvor individuelle træk ved en kompleks taktil stimulus er ekstraheret af de tre hårsækketyper og transporteret via aktiviteterne i unikke kombinationer af A? -, A? - og C-fibre til dorsalt horn.
De viste, at den genetiske mærkning af tyrosinhydroxylase-positive (TH +) DRG-neuroner karakteriserer en population af ikke-peptidergiske sensoriske neuroner med lille diameter og giver mulighed for visualisering af C-LTMR perifere ender i huden. Overraskende viste det sig, at de axoneale grene af individuelle C-LTMR'er arboriserede og dannede langsgående lansformede ender, der er tæt forbundet med zigzag (80% af enderne) og syl / auchen (20% af enderne), men ikke tylotrich hårsækkene [Fig. 1 (A4)]. Langsgående lansetformede ender har længe været anset for at høre udelukkende til A? -LTMR'er, og det var derfor uventet, at enderne af C-LTMR'er ville danne langsgående, lansettede ender.15 Disse C-LTMR'er har en mellemliggende tilpasning i sammenligning med de langsomt og hurtigt tilpasning myeliniserede mekanoreceptorer [fig. 2 (C1)].
Figur 2. Taktile receptorer i pattedyr: Kutane taktile receptorer adskiller sig til uskadelig berøring understøttet af flere receptorer med lav mekanisk tærskel (LTMR'er) i glat og håret hud og skadelig berøring understøttet af højmekanisk tærskelreceptor (HTMR'er). De udgør nervefri ender, der primært ender i epidermis. (A) Glat hud. A1: Meissner-legemer registrerer hudbevægelse og glider af genstand i hånden. De er vigtige for at aflevere objekt og fingerfærdighed. Receptorer tilpasser sig hurtigt til stimulus, er forbundet med A? fibre og tyndt til C-fibre og har stort receptorfelt. A2: Ruffini-legemer registrerer strækning af huden og er vigtige for at opdage fingerposition og afleveringsgenstand. Receptoren tilpasser sig langsomt til stimulus og opretholdt aktivitet, så længe stimulus blev anvendt. Receptorer er forbundet til A? fibre og har stort modtageligt felt. A3: Paciniske legemer er dybere i dermis og registrerer vibrationer. Receptorer er forbundet til A? fibre; de tilpasser sig hurtigt til stimulus og har det største modtagelige felt. (B) Hele huden. B1: Merkel-cellekomplekser er til stede i både glat hud og omkring hår. De er tæt udtrykt i hånden og er vigtige for teksturopfattelse og fineste diskrimination mellem to punkter. De er ansvarlige for fingerpræcision. Receptorer er forbundet til A? fibre; de tilpasser sig langsomt til stimulus og har et kort modtageligt felt. B2: skadelige berørings-HTMR'er med meget langsom tilpasning til stimulus, dvs. aktive, så længe den nociceptive stimulus anvendes. De er dannet af den frie nerveende af A? og C-fibre associeret med keratinocytter. (C) Behåret hud. C1: Hårsækkene er forbundet med de forskellige hårtyper. Hos mus er beskyttelseshår den længste og tyndt udtrykte, syl / auchenne er af mellemstørrelse, og zigzag er det mindste og tættest udtrykte hår. De er forbundet til A? fibre men også til A? og C-LTMRs fibre til syl / auchenne og zizag hår. De registrerer hårbevægelser inklusive behagelig berøring under kærtegn. De tilpasser sig hurtigt eller med mellemliggende kinetiske til stimulus. C2: C-touch nerveender svarer til en undertype af C-fibre terminal med fri slutning karakteriseret ved en lav mekanisk tærskel. De formodes at kode for behagelig fornemmelse fremkaldt af kærtegn. De tilpasser sig moderat til stimulus og har et kort modtageligt felt. Formodede mekanosensitive (MS) ionkanaler udtrykt i de forskellige taktile receptorer er angivet i overensstemmelse hermed til foreløbige data og opsummerer den nuværende hypotese under evaluering.
En anden identificeret større population vedrører A? -LTMR-enderne i Awl / Auchenne og zigzag-follikler, der skal sammenlignes med Down-hårsækken, som er grundigt undersøgt hos kat og kanin. Ginty og samarbejdspartnere viste, at TrkB udtrykkes ved høje niveauer i en delmængde af medium-diametre DRG-neuroner. Intracellulære optagelser ved hjælp af ex vivo hud-nervepræparation af mærkede fibre afslørede, at de udviser de fysiologiske egenskaber af fibre, der tidligere er undersøgt hos kat og kanin: udsøgt mekanisk følsomhed (Von Frey-tærskel <0.07 mN), der hurtigt tilpasser responser til suprathreshold stimuli, mellemledning hastigheder (5.8 0.9 m / s) og smalle ikke-bøjede soma pigge.15 Disse A? -LTMR'er danner langsgående lansetformede ender forbundet med stort set alle zigzag og syl / auchen hårsækken i bagagerummet [fig. 1 (A5)].
Endelig viste de, at de perifere ender ved hurtigt at tilpasse A? LTMR'er danner langsgående lansformede ender forbundet med vagt (eller tylotrich) og syl / auchen-hårsækkene [fig. 1 (A6)]. 15 Derudover er vagthår også forbundet med et Merkel-cellekompleks, der danner en berøringshoved forbundet med A? langsomt tilpasning af LTMR [fig. 1 (A7)].
Sammenfattende er næsten alle zigzag-hårsækkene innerveret af både C-LTMR- og Aa-LTMR-lancetformede ender; awl / auchene hår er tredobbelt innerveret af A? hurtig tilpasning-LTMR, Aa-LTMR og C-LTMR lancetformede ender; Guard hårsækkene er innerveret af A? hurtigt tilpasning-LTMR langsgående lansetformede ender og interagere med A? langsomt tilpasning-LTMR af touch dome-ender. Således modtager hver musefollikel unikke og uforanderlige kombinationer af LTMR-ender svarende til neurofysiologisk adskilte mekanosensoriske endeorganer. I betragtning af det iterative arrangement af disse tre hårtyper foreslår Ginty og samarbejdspartnere, at behåret hud består af iterativ gentagelse af perifer enhed, der indeholder, (1) et eller to centralt beliggende beskyttelseshår, (2) ~ 20 omgivende syl / auchennehår og (3 ) ~ 80 islagede zigzaghår [fig. 2 (C1)].
Rygmarv fremspring. De centrale fremskrivninger af A? hurtigt tilpasnings-LTMR'er, Aa-LTMR'er og C-LTMR'er afsluttes i distinkte, men delvist overlappende laminater (II, III, IV) i rygmarvets dorsale horn. Derudover er de centrale terminaler af LTMR'er, der innerverer de samme eller tilstødende hårsækkene i en perifer LTMR-enhed, justeret for at danne en smal LTMR-søjle i rygmarvets dorsale horn [fig. 1 (B1)]. Det ser således ud til, at en kile eller søjle af somatotopisk organiserede primære sensoriske afferente ender i dorsalhornet repræsenterer justeringen af de centrale fremspring af A? -, A? - og C-LTMR'er, der innerverer den samme perifere enhed og detekterer mekanisk stimuli, der virker på den samme lille gruppe af hårsækkene. Baseret på antallet af beskyttelses-, syl / auchen- og zigzaghår i bagagerummet og lemmerne og antallet af hver LTMR-undertype, anslår Ginty og samarbejdspartnere, at musens dorsale horn indeholder 2,000 4,000 LTMR-søjler, hvilket svarer til det omtrentlige antal perifere enheder LTMR-enheder.15
Endvidere er axoner af LTMR-undertyper tæt forbundet med hinanden med sammenflettede fremspring og interdigiterede lansetformede ender, der innerverer den samme hårsækken. Derudover, fordi de tre hårfollikeltyper udviser forskellige former, størrelser og cellulære sammensætninger, vil de sandsynligvis have tydelige afbøjnings- eller vibrationsindstillingsegenskaber. Disse fund er i overensstemmelse med klassiske neurofysiologiske målinger hos katten og kaninen, hvilket indikerer, at A? RA-LTMR'er og Aa-LTMR'er kan aktiveres differentielt ved afbøjning af forskellige hårsækketyper.16,17
Afslutningsvis er touch i håret hud kombinationen af: (1) de relative tal, unikke rumlige udbredelser og forskellige morfologiske og deflekterende egenskaber af de tre typer hårsække; (2) de unikke kombinationer af LTMR subtype-slutninger associeret med hver af de tre hårfollikeltyper; og (3) særskilte følsomheder, ledningshastigheder, spike-togmønstre og tilpasningsegenskaber af de fire hovedklasser af hårfollikelassocierede LTMR'er, som gør det hårde hudmekanosensory-system ekstraheret og overført til CNS de komplekse kombinationer af kvaliteter, som definerer en røre ved.
Free-Nerve Endings LTMRs
Generelt er C-fibre frie ender i huden HTMR'er, men en underpopulation af C-fibre reagerer ikke på skadelig berøring. Denne delmængde af taktile C-fibre (CT) afferenter repræsenterer en særskilt type umyeliniserede, lavterskel mekanoreceptive enheder, der findes i den hårede men ikke glatte hud hos mennesker og pattedyr [fig. 1 (A8)]. 18,19 CT'er er generelt forbundet med opfattelsen af behagelig taktil stimulering i kropskontakt.20,21
CT-afferenter reagerer på fordybningskræfter i området 0.3 2.5 mN og er således lige så følsomme over for huddeformation som mange af A? afferenter.19 Tilpasningskarakteristikaene for CT-afferenter er således mellemliggende i sammenligning med de langsomt og hurtigt tilpasning af myeliniserede mekanoreceptorer. De receptive felter af humane CT-afferenter er omtrent runde eller ovale i form. Feltet består af en til ni små responsive pletter fordelt over et område op til 35 mm2.22 Mushomologreceptorerne er organiseret i et mønster af diskontinuerlige pletter, der dækker ca. 50% af arealet i den hårede hud [fig. 60 (C2)]. 2
Bevis fra patienter, der mangler myelinerede taktile afferenter, indikerer, at signalering i CT-fibre aktiverer den økologiske cortex. Da dette system er dårligt ved kodning af diskriminerende aspekter ved berøring, men velegnet til kodning af langsomt og forsigtigt berøring, kan CT-fibre i hårig hud være en del af et system til behandling af behagelige og samfundsmæssige relevante aspekter af berøring. 24 CT-fiberaktivering kan også har en rolle i smertehæmning, og det er for nylig blevet foreslået, at betændelse eller traume kan ændre sensationen transporteret af C-fiber LTMR'er fra behagelig berøring til smerte. 25,26
Hvilken pathway CT-afferents rejse er endnu ikke kendt [Fig. 1 (B2)], men lavt tærskel-taktile indgange til spinothalamiske projektionsceller er blevet dokumenteret. 27-udlån anerkender rapporter om subtile kontralaterale underskud af trykdetektering hos menneskelige patienter efter ødelæggelse af disse veje efter chordotomi-procedurer. 28
LTMR i glabro hud
Merkel celle-neurit komplekser og touch dome. Merkel (1875) var den første til at give en histologisk beskrivelse af klynger af epidermale celler med store lobulerede kerner, der kom i kontakt med formodede afferente nervefibre. Han antog, at de underkendte følelse af berøring ved at kalde dem Tastzellen (taktile celler). Hos mennesker beriges Merkel-celleneuritkomplekser i berøringsfølsomme områder af huden, de findes i det basale lag af epidermis i fingre, læber og kønsorganer. De findes også i behåret hud med lavere tæthed. Merkel celle neuritkomplekset består af en Merkel celle i tæt position til en forstørret nerveterminal fra en enkelt myeliniseret A? fiber [fig. 1 (C1)] (gennemgang i Halata og samarbejdspartnere) .29 På den epidermale side udviser Merkel-celle fingerlignende processer, der strækker sig mellem nærliggende keratinocytter [fig. 1 (C2)]. Merkel-celler er keratinocyt-afledte epidermale celler.30,31 Touch touch dome blev introduceret for at navngive den store koncentration af Merkel-cellekomplekser i den hårede hud på kattepoten. En berøringskuppel kunne have op til 150 Merkel-celler, der er innerveret af en enkelt A? -Fiber og hos mennesker foruden A? -Fibre, A? og C-fibre var også regelmæssigt til stede. 32-34
Stimulering af Merkel-celleneuritkomplekser resulterer i langsomt tilpasning af Type I (SA I) -reaktioner, der stammer fra punkterede modtagelige felter med skarpe grænser. Der er ingen spontan udledning. Disse komplekser reagerer på fordybningens dybde og har den højeste rumlige opløsning (0.5 mm) af de kutane mekanoreceptorer. De transmitterer et præcist rumligt billede af taktile stimuli og foreslås at være ansvarlige for form og teksturdiskrimination [fig. 2 (B1)]. Mus blottet for Merkel-celler kan ikke registrere strukturerede overflader med deres fødder, mens de gør det ved hjælp af deres whiskers.35
Uanset om Merkel-cellen, den sensoriske neuron eller begge steder er mekanotransduktion, er det stadig et spørgsmål om debat. Hos rotter afskaffer fototoksisk ødelæggelse af Merkel-celler SA I-respons.36 Hos mus med genetisk undertrykkede Merkel-celler forsvandt SA I-svaret, der blev registreret i ex vivo hud / nervepræparat, fuldstændigt, hvilket viser, at Merkel-celler er nødvendige for korrekt kodning af Merkel receptorresponser.37 Den mekaniske stimulering af isolerede Merkel-celler i kultur ved motordrevet tryk genererer imidlertid ikke mekanisk styrede strømme.38,39 Keratinocytter kan spille en vigtig rolle i den normale funktion af Merkel-cellenneuritkomplekset. Merkel-cellefingerlignende processer kan bevæge sig med huddeformation og epidermis cellebevægelse, og dette kan være det første trin i mekanisk transduktion. Det er klart, at de betingelser, der kræves for at studere mechano-følsomhed af Merkel-celler, endnu ikke er etableret.
Ruffini slutninger. Ruffini-ender er tynde cigareformede indkapslede sensoriske ender forbundet med A? nerveender. Ruffini-ender er små bindevævscylindre arrangeret langs dermale kollagenstrenge, der leveres af en til tre myeliniserede nervefibre med en diameter på 4 mm. Op til tre cylindre med forskellig orientering i dermis kan smelte sammen og danne en receptor [fig. 6 (C1)]. Strukturelt ligner Ruffini-enderne Golgi senerorganer. De udtrykkes bredt i dermis og er blevet identificeret som de langsomt tilpasende kutane mekanoreceptorer af type II (SA II). På baggrund af spontan nervøs aktivitet fremkaldes en langsomt tilpasning af regelmæssig afladning ved vinkelret lav kraft, der opretholdes mekanisk stimulering eller mere effektivt ved hududstrækning. SA II-respons stammer fra store modtagelige felter med obskure grænser. Ruffini-receptorer bidrager til opfattelsen af retningen af objektbevægelse gennem mønsteret af hudstrækning [fig. 3 (A2)].
I mus kan SA I- og SA II-reaktioner adskilles elektrofysiologisk i ex-vivo nervehud-præparation. 40 Nandasena og samarbejdspartnere rapporterede immunokalokalisering af aquaporin 1 (AQP1) i de periodontale Ruffini-slutninger af rotteformerne, hvilket tyder på, at AQP1 er involveret i opretholdelsen af den dental osmotiske balance, der er nødvendig for mekanotransduktion. 41 De periodontale Ruffini-endinger udtrykte også den formodede mekanosensitive ionkanal ASIC3.42
Meissner-legemer. Meissner-legemer er lokaliseret i den glatte hudens dermale papiller, hovedsageligt i håndflader og fodsåler, men også i læber, tunge, ansigt, brystvorter og kønsorganer. Anatomisk består de af en indkapslet nerveende, hvor kapslen er lavet af flade understøttende celler arrangeret som vandrette lameller indlejret i bindevæv. Der er en enkelt nervefiber A? afferenter forbundne pr. legemsvæske [Fig. 1 (C4)]. Enhver fysisk deformation af kroppen viser en volley af handlingspotentialer, der hurtigt ophører, dvs. de tilpasser sig hurtigt receptorer. Når stimulus fjernes, genvinder kroppsformen sin form og producerer samtidig en ny volley af handlingspotentialer. På grund af deres overfladiske placering i dermis reagerer disse kroppe selektivt på hudbevægelse, taktil detektion af glidning og vibrationer (20 Hz). De er følsomme over for dynamisk hud - for eksempel mellem huden og en genstand, der håndteres [fig. 40 (A2)].
Pacinian corpuscles. Pacininske legemer er de dybere mekanoreceptorer i huden og er den mest følsomme indkapslede kutane mekanoreceptor for hudbevægelse. Disse store ovale blodlegemer (1 mm i længden) lavet af koncentriske lameller af fibrøst bindevæv og fibroblaster foret med fladt modificerede Schwann-celler udtrykkes i den dybe dermis.43 I midten af kroppen, i et væskefyldt hulrum kaldet indre pære , afslutter en enkelt A? afferent umyeliniseret nerveende [fig. 1 (C5)]. De har et stort modtageligt felt på hudens overflade med et særligt følsomt center. Udviklingen og funktionen af flere hurtigt tilpassende mekanoreceptortyper forstyrres i c-Maf-mutante mus. Især Pacininske kroppe er alvorligt atrofiere.44
Pacininske blodlegemer viser meget hurtig tilpasning som reaktion på fordybningen af huden, den hurtigt tilpasende II (RA II) nerveafladning, der er i stand til at følge høj frekvens af vibrerende stimuli, og tillader opfattelse af fjerne begivenheder gennem transmitterede vibrationer.45 Pacinian corpuscle afferenter reagerer på vedvarende fordybning med forbigående aktivitet ved stimulusens begyndelse og forskydning. De kaldes også accelerationsdetektorer, fordi de kan registrere ændringer i stimulusens styrke, og hvis ændringshastigheden i stimulus ændres (som det sker i vibrationer), bliver deres svar proportionalt med denne ændring. Paciniske legemer mærker grove trykændringer og mest af alt vibrationer (150 300 Hz), som de kan registrere selv centimeter væk [fig. 2 (A3)].
Tonisk respons blev observeret i decapsulated Pacinian corpuscle.46 Derudover reagerer intakte Pacinian-korpusker med vedvarende aktivitet under konstante indrykkestimuli uden at ændre mekaniske tærskler eller responsfrekvens, når GABA-medieret signalering er blokeret mellem lamellatglia og en nerveendring. 47. de ikke-neuronale komponenter i Pacinian-korpuslet kan have dobbelt roller i filtrering af den mekaniske stimulus såvel som til modulering af sensory neurons responsegenskaber.
Rygmarv fremspring. Fremskrivninger af A? -LTMR'erne i rygmarven er opdelt i to grene. Den vigtigste centrale gren stiger op i rygmarven i de ipsilaterale dorsale søjler til cervikal niveau [fig. 1 (B3)]. Sekundære grene slutter i ryghornet i laminerne IV og interfererer f.eks. Med smerteoverførslen. Dette kan dæmpe smerter som en del af portkontrollen [Fig. 1 (B4)]. 48
Ved cervicale niveauer separerer hovedpartens axoner i to områder: Midterbanen omfatter gracile fasciklen, der formidler information fra den nedre halvdel af kroppen (ben og stammen), og den ydre kanal omfatter den cuneate fascicle, der transporterer information fra den øvre halvdel af kroppen (arme og kuffert) [Fig. 1 (B5)].
Primære taktile afferenter laver deres første synaps med anden ordens neuroner ved medulla, hvor fibre fra hver kanal synapser i en kerne med samme navn: gracile fasciculus axones synaps i gracile nucleus og cuneat axones synapse i cuneatkernen [Fig. 1 (B6)]. Neuroner, der modtager synapsen, tilvejebringer de sekundære afferenter og krydser midtlinjen straks for at danne en kanal på den kontralaterale side af hjernestammen den mediale lemniscus som stiger gennem hjernestammen til den næste relæstation i midthjernen, specifikt i thalamus [Fig. . 1 (B7)].
Molekylær specifikation af LTMR'er. Molekylære mekanismer, der styrer den tidlige diversificering af LTMR'er, er for nylig blevet delvist belyst. Bourane og samarbejdspartnere har vist, at de neuronale populationer, der udtrykker Ret tyrosinkinasereceptoren (Ret) og dens co-receptor GFR? 2 i E11 13 embryonale mus, DRG selektivt udtrykker transkriptionsfaktoren Mafa.49,50 Disse forfattere viser, at Mafa / Ret / GFR2-neuroner bestemt til at blive tre specifikke typer LTRM'er ved fødslen: SA1-neuroner, der innerverer Merkel-cellekomplekser, de hurtigt tilpasende neuroner, der innerverer Meissner-legemer og de hurtigt tilpasende afferenter (RA I), der danner lancetformede ender omkring hårsækkene. Ginty og samarbejdspartnere rapporterer også, at DRG-neuroner, der udtrykker early-Ret, hurtigt tilpasser sig mekanoreceptorer fra Meissner-blodlegemer, Pacinian-blodlegemer og lancetformede ender omkring hårsækkene.51 De innerverer diskrete målzoner inden for gracile og cuneate kerner og afslører et modalitetsspecifikt mønster af mekanosensor neuronale aksonale fremspring i hjernestammen.
Undersøgelse af humane hudmekanoreceptorer. Teknikken til mikroneurografi beskrevet af Hagbarth og Vallbo i 1968 er blevet anvendt til at undersøge afladningsadfærden for enkelt menneskelige mekaniske følsomme ender, der forsyner muskel, led og hud (se til gennemgang Macefield, 2005). 52,53 Størstedelen af mikronurografi på huden undersøgelser har karakteriseret fysiologien af taktile afferenter i håndens glat hud. Mikroelektrodeoptagelser fra median- og ulnarnervene hos mennesker har afsløret berøringsfølelse genereret af de fire klasser af LTMR'er: Meissner-afferenter er særligt følsomme over for lysstrøg over huden og reagerer på lokale forskydningskræfter og begyndende eller åbenlyse glider inden i det modtagelige felt. Paciniske afferenter er udsøgte følsomme over for hurtige mekaniske transienter. Tilbudere reagerer kraftigt på at blæse over det modtagelige felt. Et Pacinian-legeme placeret i et ciffer vil normalt svare på at banke på bordet, der understøtter armen. Merkel-afferenter har karakteristisk en høj dynamisk følsomhed over for indrykningsstimuli anvendt på et diskret område og reagerer ofte med en afladning under frigivelse. Selvom Ruffini-afferenterne reagerer på kræfter, der påføres normalt på huden, er et unikt træk ved SA II-afferenter deres evne til at reagere også på lateral hudstrækning. Endelig har hårenheder i underarmen store ovale eller uregelmæssige modtagelige felter sammensat af flere følsomme pletter, der svarede til individuelle hår (hver afferent forsyning ~ 20 hår).
Keratinocytternes mekaniske følsomhed
Enhver mekanisk stimulering på huden skal overføres gennem keratinocytter, der danner epidermis. Disse allestedsnærværende celler kan udføre signaleringsfunktioner ud over deres støttende eller beskyttende roller. For eksempel udskiller keratinocytter ATP, et vigtigt sensorisk signalmolekyle som reaktion på mekaniske og osmotiske stimuli. 54,55 Udløsningen af ATP inducerer intracellulær calciumforøgelse ved autokrin stimulering af purinergreceptorer. 55 Derudover er der tegn på, at hypotonicitet aktiverer Rho-kinasen signalveje og den efterfølgende F-actin-spændingsfiberdannelse, hvilket tyder på, at den mekaniske deformation af keratinocytterne mekanisk kan interferere med nabo-cellerne, såsom Merkel-celler, til uskadelig berøring og C-fiberfri ende for skadelig berøring [Fig. 1 (C6)]. 56,57
Skadelig berøring
Høj tærskel mekanoreceptorer (HTMR'er) er epidermale C- og A? gratis nerveender. De er ikke forbundet med specialiserede strukturer og observeres i begge hårhud [fig. 1 (A9)] og glat hud [fig. 1 (C7)]. Imidlertid skal betegnelsen fri nerveendende betragtes med forsigtighed, da nerveender altid er i tæt placering med keratinocyt- eller Langherans-celle eller melanocytter. Ultrastrukturel analyse af nerveender afslører tilstedeværelsen af groft endoplasmatisk retikulum, rigelig mitokondrier og tætkerne-vesikel. Tilstødende membraner af epidermale celler fortykkes og ligner postsynaptisk membran i nervevæv. Bemærk, at interaktionerne mellem nerveender og epidermale celler kan være tovejs, da epidermale celler kan frigive mediatorer som ATP, interleukin (IL6, IL10) og bradykinin, og omvendt kan peptidergiske nerveender frigive peptider såsom CGRP eller substans P, der virker på epidermale celler. HTMR'er omfatter mechano-nociceptorer, der kun ophidses af skadelige mekaniske stimuli og polymodale nociceptorer, der også reagerer på skadelig varme og eksogent kemikalie [Fig. 2 (B2)] 58
HTMR-afferente fibre slutter på projektionsneuroner i rygmarvs dorsale horn. A? -HTMR'er kommer i kontakt med andenordens neuroner overvejende i lamina I og V, hvorimod C-HTMR'er afsluttes i lamina II [Fig. 1 (B8)]. Anden ordens nociceptive neuroner rager ud til den kontrolsidede side af rygmarven og stiger op i det hvide stof og danner det anterolaterale system. Disse neuroner slutter hovedsageligt i thalamus [fig. 1 (B9 og B10)].
Mechano-strømme i Somatosensoriske Neuroner
Mekanismerne til langsom eller hurtig tilpasning af mekanoreceptorer er endnu ikke klarlagt. Det er uklart, i hvilket omfang mekanoreceptortilpasning tilvejebringes af den cellulære miljø i den sensoriske nerveafslutning, de mekaniske gatedkanalers egenskaber og egenskaberne af de aksonale spændingsgatede ionkanaler i sensoriske neuroner (Fig. 2). De seneste fremskridt i karakteriseringen af mekanisk gatedstrømme har imidlertid vist, at forskellige klasser af mekanosensitive kanaler findes i DRG-neuroner og kan forklare nogle aspekter af tilpasningen af mekanoreceptorer.
In vitro-registrering hos gnavere har vist, at somaen fra DRG-neuroner er iboende mekanosensitiv og udtrykker kationiske mechano-gatede strømme.59-64 Gadolinium og rutheniumrød blokerer fuldt ud mekanosensitive strømme, hvorimod eksternt calcium og magnesium ved fysiologiske koncentrationer samt amilorid og benzamil, forårsager delvis blokering. 60,62,63 FM1-43 fungerer som en varig blokering, og injektionen af FM1-43 i musens bagpote nedsætter smertefølsomheden i Randall Selitto-testen og øger den vurderede tærskel for tilbagetrækning af poten med von Frey-hår.65
Som reaktion på vedvarende mekanisk stimulering falder mekanosensitive strømme gennem lukning. Baseret på tidskonstanterne for det nuværende forfald er der adskilt fire forskellige typer af mekanosensitive strømme: hurtigt tilpasning af strømme (~ 3 6 ms), mellemliggende tilpasning af strømme (~ 15 30 ms), langsomt tilpasning af strømme (~ 200 300 ms) ) og ultra-langsomt tilpasningsstrømme (~ 1000 ms) .64 Alle disse strømme er til stede med variabel forekomst i rotte-DRG-neuroner, der innerverer bagbenets glat hud.64
Den mekaniske følsomhed af mekanosensitive strømme kan bestemmes ved at anvende en række inkrementelle mekaniske stimuli, hvilket giver mulighed for relativt detaljeret stimulusstrømsanalyse.66 Stimulus strømsforholdet er typisk sigmoidalt, og den maksimale amplitude af strømmen bestemmes af antallet af kanaler, der er åbne samtidigt.64,67 Interessant nok er det blevet rapporteret, at den hurtigt tilpasende mekanosensitive strøm viser lav mekanisk tærskel og halvaktiverende midtpunkt sammenlignet med den ultra-langsomt tilpasende mekanosensitive strøm.63,65
Sensoriske neuroner med ikke-nociceptive fænotyper udtrykker fortrinsvis hurtig tilpasning af mekanosensitive strømme med lavere mekanisk tærskel. 60,61,63,64,68 Omvendt rapporteres langsomt og ultra-langsomt tilpasning af mekanosensitive strømme i formodede ikke-nociceptive celler. 64,68 Dette fremkaldte forslag om, at disse strømme kunne bidrage til forskellige mekaniske tærskler ses i LTMR'er og HTMR'er in vivo. Skønt disse in vitro-forsøg bør tages med forsigtighed, blev der også tilvejebragt støtte til tilstedeværelsen i DR-neuronernes neuroner af lav- og høj-tærskelmekanotransducere ved radial strækbaseret stimulation af dyrkede mus-sensoriske neuroner. 69 Dette paradigme afslørede to hovedpopulationer af strækningskänslige neuroner, en som reagerer på lav stimulus amplitude og en anden som selektivt reagerer på høj stimulus amplitude.
Disse resultater har vigtige, men spekulative, mekaniske implikationer: Den mekaniske tærskel af sensoriske neuroner kan have lidt at gøre med den mekaniske organisering af mekanoreceptoren, men kan ligge i egenskaberne af de mekanisk gatede ionkanaler.
Mekanismerne, der ligger til grund for desensibilisering af mekanosensitive kationstrømme i DRG-neuroner fra rotter, er for nylig blevet afsløret.64,67 Det skyldes to samtidige mekanismer, der påvirker kanalegenskaber: tilpasning og inaktivering. Tilpasning blev først rapporteret i auditive hårcelleundersøgelser. Det kan beskrives operationelt som en simpel oversættelse af transducerkanalens aktiveringskurve langs den mekaniske stimulusakse. 70-72 Tilpasning tillader sensoriske receptorer at opretholde deres følsomhed over for nye stimuli i nærværelse af en eksisterende stimulus. Imidlertid kan en væsentlig del af mekanosensitive strømme i DRG-neuroner ikke genaktiveres efter konditionering af mekanisk stimulering, hvilket indikerer inaktivering af nogle transducerkanaler.64,67 Derfor virker både inaktivering og tilpasning i tandem for at regulere mekanosensitive strømme. Disse to mekanismer er fælles for alle mekanosensitive strømme identificeret i rotte-DRG-neuroner, hvilket tyder på, at relaterede fysisk-kemiske elementer bestemmer kinetikken for disse kanaler.64
Afslutningsvis er det afgørende at bestemme egenskaberne af endogene mekanosensitive strømme in vitro i søgen efter at identificere transduktionsmekanismer på molekylært niveau. Variationen observeret i den mekaniske tærskel og tilpasningskinetikken for de forskellige mekanisk gatede strømme i DRG-neuroner antyder, at ionkanalernes indre egenskaber i det mindste delvist kan forklare mekanisk tærskel- og tilpasningskinetik hos de mekanoreceptorer, der er beskrevet i årtierne 1960 80 ved anvendelse af ex vivo-præparater.
Putative Mekanosensitive Proteiner
Mekanisk følsomme ionstrømme i somatosensoriske neuroner karakteriseres godt, derimod er der ikke meget kendt om identiteten af molekyler, som medierer mekanotransduktion hos pattedyr. Genetiske skærme i Drosophila og C. elegans har identificeret kandidatmekanotransduktionsmolekyler, herunder TRP- og degenerin / epithelial Na + -kanalen (Deg / ENaC) -familierne. 73 Nylige forsøg på at belyse molekylære grundlag for mekanotransduktion hos pattedyr har i høj grad fokuseret på homologer af disse kandidater . Derudover er mange af disse kandidater til stede i kutane mekanoreceptorer og somatosensoriske neuroner (Fig. 2).
Acid-Sensing Ion Channels
ASIC'er tilhører en proton-gated undergruppe af degenerin epithelial Na + kanalfamilien.74 Tre medlemmer af ASIC-familien (ASIC1, ASIC2 og ASIC3) udtrykkes i mekanoreceptorer og nociceptorer. ASIC-kanalers rolle er blevet undersøgt i adfærdsmæssige studier ved anvendelse af mus med målrettet sletning af ASIC-kanalgener. Sletning af ASIC1 ændrer ikke funktionen af kutane mekaniske receptorer, men øger afferenters mekaniske følsomhed, der innerverer tarmen.75 ASIC2 knockout-mus udviser en nedsat følsomhed for hurtigt tilpasning af kutane LTMR'er.76 Imidlertid rapporterede efterfølgende studier manglende virkninger af at slå ASIC2 ud på både visceral mechano-nociception og kutan mekanosensation.77 ASIC3-forstyrrelse mindsker mechano-følsomhed hos viscerale afferenter og reducerer responsen af kutane HTMR'er på skadelige stimuli.76
Den Transient Receptor Channel
TRP superfamilien er opdelt i seks subfamilier i pattedyr. 78 Næsten alle TRP-underfamilier har medlemmer knyttet til mekanosensation i en række cellesystemer. 79 I pattedyrs sensoriske neuroner er TRP-kanaler imidlertid bedst kendt for at detektere termisk information og formidlende neurogen inflammation, og kun to TRP-kanaler, TRPV4 og TRPA1, har været impliceret i kontaktfølsomhed. Forstyrrende TRPV4-ekspression hos mus har kun beskedne virkninger på akutte mekanosensoriske tærskler, men reducerer kraftigt følsomheden overfor skadelige mekaniske stimuli. 80,81 TRPV4 er en afgørende determinant i formgivning af nociceptive neurons reaktion på osmotisk stress og mekanisk hyperalgesi under betændelse. 82,83 TRPA1 synes at har en rolle i mekanisk hyperalgesi. TRPA1-mangelfulde mus udviser smerteoverfølsomhed. TRPA1 bidrager til transduktion af mekaniske, kolde og kemiske stimuli i nociceptor sensoriske neuroner, men det ser ud til, at det ikke er vigtigt for hårcelletransduktion. 84,85
Der er intet klart bevis på, at TRP-kanaler og ASIC-kanaler, der er udtrykt i pattedyr, er mekanisk gatede. Ingen af disse kanaler udtrykker heterologt rekapitulerer den elektriske signatur af mekanosensitive strømme observeret i deres native miljø. Dette udelukker ikke muligheden for, at ASIC'er og TRPs-kanaler er mekanotransducere, givet usikkerheden om, hvorvidt en mekanotransduktionskanal kan fungere uden for sin cellulære kontekst (se afsnit om SLP3).
Piezo Proteiner
Piezo protiens er for nylig blevet identificeret som lovende kandidater til mekanosenserende proteiner fra Coste og collaborators.86,87 Vertebrates har to Piezo-medlemmer, Piezo 1 og Piezo 2, der tidligere er kendt som FAM38A og FAM38B, som er godt konserveret gennem multi-cellulære eukaryoter. Piezo 2 er rigeligt i DRG'er, mens Piezo 1 næppe kan påvises. Piezo-inducerede mekanosensitive strømme forhindres hæmmet af gadolinium, rutheniumrød og GsMTx4 (et toksin fra tarantula Grammostola spatulata) .88 Ekspression af Piezo 1 eller Piezo 2 i heterologe systemer frembringer mekanosensitive strømme, kinetikken ved inaktivering af Piezo 2 strøm er hurtigere end Piezo 1. På samme måde som endogene mekanosensitive strømme har Piezo-afhængige strømme reverseringspotentialer omkring 0 mV og er kation-ingen selektive, med Na +, K +, Ca2 + og Mg2 +, der gennemsyrer den underliggende kanal. På samme måde reguleres piezoafhængige strømme af membranpotentiale med en markant bremsning af den nuværende kinetik ved depolariserede potentialer. 86
Piezo proteiner er utvivlsomt mekanosenserende proteiner og deler mange egenskaber ved hurtigt at tilpasse mekanosensitive strømme i sensoriske neuroner. Behandling af dyrkede DRG-neuroner med Piezo 2-kort interfererende RNA reducerede andelen af neuroner med hurtig tilpasning af strøm og nedsat procentdelen af mekanosensitive neuroner. 86 Transmembrane-domæner er placeret i hele piezo-proteinerne, men der er ingen indlysende poreholdige motiver eller ionkanal signaturer blevet identificeret. Mus Piezo 1-protein renset og rekonstitueret i asymmetriske lipid-bilayere og liposomer danner ionkanaler, der er følsomme for rutheniumrød. 87 Et vigtigt trin i validerende mekanotransduktion gennem Piezo-kanaler er at anvende in vivo-fremgangsmåder til at bestemme den funktionelle betydning i berøringssignalering. Oplysninger blev givet i Drosophila, hvor sletning af det enkelte Piezo-medlem reducerede mekanisk reaktion på skadelige stimuli uden at påvirke normal berøring. 89 Selvom deres struktur fortsat skal bestemmes, er denne nye familie af mekanosensitive proteiner et lovende emne til fremtidig forskning ud over grænsen af touch sensation. For eksempel viser en nylig undersøgelse af patienter med anæmi (arvelig xerocytose) Piezo 1's rolle ved opretholdelse af erytrocytvolumen homeostase.90
Transmembrane Channel-Like (TMC)
En nylig undersøgelse viser, at to proteiner, TMC1 og TMC2 er nødvendige for hårcellemekanotransduktion. 91 Arvelig døvhed på grund af TMC1-genmutation blev rapporteret hos mennesker og mus. 92,93 Tilstedeværelsen af disse kanaler var endnu ikke vist i det somatosensoriske system, men det ser ud til at være en god ledelse til at undersøge.
Stomatin-lignende Protein 3 (SLP3)
Ud over transduktionskanalerne har nogle tilbehørsproteiner forbundet med kanalen vist sig at spille en rolle i berøringsfølsomhed. SLP3 udtrykkes i mammale DRG neuroner. Undersøgelser, der anvendte mutante mus, der manglede SLP3, havde vist ændring i mekanosensation og mekanosensorrige strømme. 94,95 SLP3 præcise funktion forbliver ukendt. Det kan være en linker mellem den mekanosensitive kanal og de underliggende mikrotubuli, som foreslået for sin C. elegans homolog MEC2.96 Nyligt GR. Lewin lab har antydet, at en tether syntetiseres af DRG sensoriske neuroner og forbinder mekanosensitiv ionkanal med den ekstracellulære matrix. 97 Forstyrrende forbindelsen afskaffer den RA-mekanosensitive strøm, hvilket tyder på, at nogle ionkanaler kun er mekanisk følsomme, når de er tæmmet. RA-mekanosensitive strømme hæmmes også af laminin-332, et matrixprotein produceret af keratinocytter, der forstærker hypotesen af en modulering af den mekanosensitive strøm af ekstracellulære proteiner. 98
K + Kanalsubfamilie
Parallelt med kationiske depolariserende mekanosensitive strømme undersøges tilstedeværelsen af repolariserende mekanosensitive K + strømme. K + -kanaler i mekanosensitive celler kan træde i den aktuelle balance og bidrage til at definere den mekaniske tærskel og tidskurset for tilpasning af mekanoreceptorer.
KCNK medlemmer tilhører de to-pore domæne K + -kanal (K2P) family.99,100 The K2P vise en bemærkelsesværdig række regulering af cellulære, fysiske og farmakologiske midler, herunder pH-ændringer, varme, stræk og membran deformation. Disse K2P er aktive ved hvilemembranpotentiale. Adskillige KCNK underenheder udtrykkes i somatosensoriske neurones.101 KCNK2 (TREK-1), KCNK4 (TRAAK) og TREK-2 kanaler er blandt de få kanaler, som en direkte mekanisk gating ved membranfiltrering strækning har været shown.102,103
Mus med et forstyrret KCNK2-gen viste en forbedret følsomhed over for varme og milde mekaniske stimuli, men en normal tilbagetrækningstærskel for skadeligt mekanisk tryk på bagpoten ved anvendelse af Randall Selitto-testen.104 KCNK2-mangelfulde mus viser også øget termisk og mekanisk hyperalgesi ved inflammatorisk betingelser. KCNK4-knockout-mus var overfølsomme over for mild mekanisk stimulering, og denne overfølsomhed blev øget ved yderligere inaktivering af KCNK2.105 Øget mekanosensitivitet af disse knockout-mus kunne betyde, at stretch normalt aktiverer både depolarisering og repolarisering af mekanosensitive strømme på en koordineret måde, ligesom ubalancen mellem depolarisering og repolarisering af spændingsstyrede strømme.
KCNK18 (TRESK) er en vigtig bidragyder til baggrunden K + konduktans, der regulerer hvilemembranpotentialet for somatosensoriske neuroner.106 Selvom det ikke vides, om KCNK18 er direkte følsom over for mekanisk stimulering, kan det spille en rolle i formidling af reaktioner på let berøring, såvel som smertefulde mekaniske stimuli. KCNK18 og i mindre grad KCNK3 foreslås at være det molekylære mål for hydroxy -? - sanshool, en forbindelse fundet i Schezuan peberkorn, der aktiverer berøringsreceptorer og inducerer en prikkende fornemmelse hos mennesker. 107,108
Den spændingsafhængige K + kanal KCNQ4 (Kv7.4) er afgørende for at indstille hastigheden og frekvenspræferensen af en subpopulation af hurtigt tilpassende mekanoreceptorer i både mus og mennesker. Mutation af KCNQ4 er i første omgang forbundet med en form for arvelig døvhed. Interessant nok lokaliserer en nylig undersøgelse KCNQ4 i perifere nerveender af kutan hurtigt tilpasning af hårfollikel og Meissner corpuscle. Følgelig fører tab af KCNQ4-funktion til en selektiv forstærkning af mekanoreceptorfølsomhed over for lavfrekvent vibration. Navnlig viser personer med forsinket høretab på grund af dominerende mutationer i KCNQ4-genet forbedret ydeevne ved detektering af små amplitude, lavfrekvente vibrationer. 109
Dr. Alex Jimenez's Insight
Touch anses for at være en af de mest komplekse sanser i menneskekroppen, især fordi der ikke er noget specifikt organ der har ansvaret for det. I stedet opstår følelsesfølsomheden gennem sensoriske receptorer, kendt som mekanoreceptorer, som findes over huden og reagerer på mekanisk tryk eller forvrængning. Der findes fire hovedtyper af mekanoreceptorer i den glabroøse eller hårløse hud hos pattedyr: lamellære legemer, taktile legemer, Merkel-nerveendinger og pærekroppe. Mekanoreceptorer fungerer for at muliggøre detektion af berøring for at overvåge muskler, knogler og ledders position, kendt som proprioception, og endda at opdage lyde og bevægelse af kroppen. Forståelse af mekanismerne for struktur og funktion af disse mekanoreceptorer er et grundlæggende element i udnyttelsen af behandlinger og terapier til smertebehandling.
Konklusion
Touch er en kompleks forstand, fordi den repræsenterer forskellige taktile kvaliteter, nemlig vibrationer, form, tekstur, fornøjelse og smerte, med forskellige diskriminerende forestillinger. Indtil videre var korrespondancen mellem et berøringsorgan og den psykofysiske sans korrelativ, og klassespecifikke molekylmarkører er bare ved at komme frem. Udviklingen af gnaverprøver, der matcher mangfoldigheden af berøringsadfærd, er nu påkrævet for at lette fremtidig genomforskning. Brugen af mus, der mangler specifikke delmængder af sensoriske afferente typer, vil i høj grad lette identifikation af mekanoreceptorer og sensoriske afferente fibre forbundet med en bestemt berøringsmodalitet. Interessant nok åbner et nyt papir det vigtige spørgsmål om det genetiske grundlag for mekanosensoriske træk hos mennesker og foreslår, at enkeltgenmutation kan påvirke berøringsfølsomheden negativt. 110 Dette understreger, at patofysiologien af det menneskelige berøringsunderskud i vid udstrækning er ukendt og ville helt sikkert fremskridt ved at identificere nøjagtigt delmængden af sensoriske neuroner forbundet med en berøringsmodalitet eller et trykunderskud.
Til gengæld er der gjort fremskridt med at definere de biokysiske egenskaber ved de mechano-gated strømme. 64 Udviklingen af nye teknikker i de senere år, som muliggør overvågning af membranspændingsændringer, mens man registrerer mechano-gated strøm, har vist sig at være en værdifuld eksperimentel metode til at beskrive mekanosensitive strømme med hurtig, mellemliggende og langsom tilpasning (gennemgået i Delmas og samarbejdspartnere) .66,111 Fremtiden vil være at bestemme de nuværende egenskabers rolle i mekanismerne til tilpasning af funktionelt forskellige mekanoreceptorer og bidraget fra mekanosensitive K + strømme til excitabiliteten af LTMR'er og HTMR'er.
Den molekylære karakter af mechano-gated strømme i pattedyr er også et fremtidigt lovende forskningsemne. Fremtidig forskning vil fremskridt i to perspektiver, først for at bestemme rollen som tilbehørsmolekyle, der tinder kanaler til cytoskelettet og ville være forpligtet til at overføre eller regulere mekanosensitivitet af ionkanaler af lignende af TRP- og ASIC / EnaC-familier. For det andet at undersøge det store og lovende område af Piezo-kanalernes bidrag ved at besvare nøglespørgsmål i forhold til permeations- og gatingmekanismerne, delmængden af sensoriske neuroner og touchmodaliteter involverer Piezo og Piezo's rolle i ikke-neuronale celler associeret med mechanosensation.
Berøringssansen i sammenligning med synet, smagen, lyden og lugten, som bruger bestemte organer til at behandle disse fornemmelser, kan forekomme overalt i kroppen gennem små receptorer kendt som mekanoreceptorer. Forskellige typer mekanoreceptorer kan findes i forskellige lag af huden, hvor de kan opdage en bred vifte af mekanisk stimulering. Artiklen ovenfor beskriver specifikke højdepunkter, der demonstrerer fremskridt med strukturelle og funktionelle mekanismer hos mekanoreceptorer forbundet med berøringssansen. Information der henvises til fra National Center for Biotechnology Information (NCBI). Omfanget af vores information er begrænset til kiropraktik såvel som til rygmarvsskader og tilstande. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emner: Rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og savnede dage på arbejdspladsen over hele verden. Faktisk er rygsmerter blevet tilskrevet som den næst mest almindelige årsag til doktorkontorbesøg, der kun overstiger luftvejsinfektioner. Ca. 80 procent af befolkningen vil opleve en form for rygsmerter mindst én gang i hele deres liv. Ryggraden er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette skader og / eller forværrede forhold, som f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
1. Moriwaki K, Yuge O. Topografiske træk ved kutane taktile hypoæstetiske og hyperæstetiske abnormiteter ved kronisk smerte.�Smerter.�1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Cross Ref]
2. Shim B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Mekanisk og varmesensibilisering af kutane nociceptorer hos rotter med eksperimentel perifer neuropati.�Neurovidenskab.�2005;132:193�201. doi: 10.1016/j.neuroscience.2004.12.036.�[PubMed]�[Cross Ref]
3. Klegetveit IP, J�rum E. Stor og lille fiberdysfunktion ved perifere nerveskader med eller uden spontan smerte.�J Pain. 2010;11:1305�10. doi: 10.1016/j.jpain.2010.03.004.�[PubMed]�[Cross Ref]
4. Noback CR. Morfologi og fylogeni af hår.�Ann NY Acad Sci.�1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed]�[Cross Ref]
5. Straile WE. Atypiske beskyttelses-hårsække i huden på kanin.�Naturen.�1958;181:1604�5. doi: 10.1038/1811604a0.�[PubMed]�[Cross Ref]
6. Straile WE. Morfologien af tylotrich follikler i huden på kanin.�Am J Anat.�1961;109:1�13. doi: 10.1002/aja.1001090102.�[PubMed]�[Cross Ref]
7. Millard CL, Woolf CJ. Sensorisk innervation af hårene på rottebagbenet: en lysmikroskopisk analyse.�J Comp Neurol.�1988;277:183�94. doi: 10.1002/cne.902770203.�[PubMed]�[Cross Ref]
9. Brown AG, Iggo A. En kvantitativ undersøgelse af kutane receptorer og afferente fibre hos katte og kaniner.�J Physiol.�1967;193:707 33. [PMC gratis artikel]�[PubMed]
10. Burgess PR, Petit D, Warren RM. Receptortyper i kattehåret hud forsynet af myelinerede fibre.�J Neurophysiol.�1968;31:833 48. [PubMed]
12. Hussein MA. Det overordnede mønster af hårsækkenes arrangement hos rotter og mus.�J Anat.�1971;109:307 16. [PMC gratis artikel]�[PubMed]
13. Vielkind U, Hardy MH. Ændring af mønstre af celleadhæsionsmolekyler under udvikling af hårsækkene fra musepelage. 2. Follikelmorfogenese i hårmutanterne, Tabby og dunet.�Acta Anat (Basel) �1996;157:183�94. doi: 10.1159/000147880.�[PubMed]�[Cross Ref]
14. Hardy MH, Vielkind U. Ændring af mønstre af celleadhæsionsmolekyler under udvikling af hårsække på musepelage. 1. Follikelmorfogenese i vildtype mus.�Acta Anat (Basel) �1996;157:169�82. doi: 10.1159/000147879.�[PubMed]�[Cross Ref]
15. Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S, et al. Den funktionelle organisation af kutane lavtærskelmekanosensoriske neuroner.�Celle.�2011;147:1615�27. doi: 10.1016/j.cell.2011.11.027.[PMC gratis artikel]�[PubMed]�[Cross Ref]
16. Brown AG, Iggo A. En kvantitativ undersøgelse af kutane receptorer og afferente fibre hos katte og kaniner.�J Physiol.�1967;193:707 33. [PMC gratis artikel]�[PubMed]
17. Burgess PR, Petit D, Warren RM. Receptortyper i kattehåret hud forsynet af myelinerede fibre.�J Neurophysiol.�1968;31:833 48. [PubMed]
18. Vallbo A, Olausson H, Wessberg J, Norrsell U. Et system af umyelinerede afferenter til uskadelig mekanoreception i den menneskelige hud.�Hjerne res.�1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed]�[Cross Ref]
19. Vallbo AB, Olausson H, Wessberg J. Umyelinerede afferenter udgør et andet system, der koder taktile stimuli af den menneskelige behårede hud.�J Neurophysiol.�1999;81:2753 63. [PubMed]
22. Wessberg J, Olausson H, Fernstr�m KW, Vallbo AB. Receptive feltegenskaber af umyelinerede taktile afferenter i den menneskelige hud.�J Neurophysiol.�2003;89:1567�75. doi: 10.1152/jn.00256.2002.�[PubMed][Cross Ref]
23. Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Molekylær genetisk visualisering af en sjælden undergruppe af umyelinerede sensoriske neuroner, der kan detektere blid berøring.�Nat Neurosci.�2007;10:946�8. doi: 10.1038/nn1937.�[PubMed]�[Cross Ref]
24. Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G, et al. Umyeliniserede taktile afferenter signalerer berøring og projicerer til den øde cortex.�Nat Neurosci.�2002;5:900�4. doi: 10.1038/nn896.[PubMed]�[Cross Ref]
25. Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. Neurofysiologien af umyeliniserede taktile afferenter.�Neurosci Biobehav Rev.�2010;34:185�91. doi: 10.1016/j.neubiorev.2008.09.011.�[Anmeldelse][PubMed]�[Cross Ref]
26. Kr�mer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M, et al. Aktivering af det kortikale smertenetværk ved blød taktil stimulering efter injektion af sumatriptan.�Smerter.�2007;133:72�8. doi: 10.1016/j.pain.2007.03.001.�[PubMed]�[Cross Ref]
27. Applebaum AE, Beall JE, Foreman RD, Willis WD. Organisation og receptive felter af primats spinothalamiske traktus neuroner.�J Neurophysiol.�1975;38:572 86. [PubMed]
28. Hvid JC, Sweet WH. Effektiviteten af chordotomi ved fantomsmerter efter amputation.�AMA Arch Neurol Psychiatry.�1952;67:315 22. [PubMed]
29. Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel og hans �Merkel-celle�, morfologi, udvikling og fysiologi: gennemgang og nye resultater.�Anat Rec A Discov Mol Cell Evol Biol.�2003;271:225�39. doi: 10.1002/ar.a.10029.�[PubMed]�[Cross Ref]
30. Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Pattedyrs Merkel-celler nedstammer fra den epidermale afstamning.�Dev Biol.�2009;336:76�83. doi: 10.1016/j.ydbio.2009.09.032.[PMC gratis artikel]�[PubMed]�[Cross Ref]
31. Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N, et al. Epidermale progenitorer giver anledning til Merkel-celler under embryonal udvikling og voksen homeostase.�J Cell Biol.�2009;187:91�100. doi: 10.1083/jcb.200907080.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
32. Ebara S, Kumamoto K, Baumann KI, Halata Z. Tredimensionelle analyser af berøringskupler i kattepotens behårede hud afslører morfologiske substrater til kompleks sensorisk behandling.�Neurosci Res.�2008;61:159�71. doi: 10.1016/j.neures.2008.02.004.�[PubMed]�[Cross Ref]
33. Guinard D, Usson Y, Guillermet C, Saxod R. Merkel komplekser af menneskelig digital hud: tredimensionel billeddannelse med konfokal lasermikroskopi og dobbelt immunfluorescens.�J Comp Neurol.�1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Cross Ref]
34. Reinisch CM, Tschachler E. Berøringskuplen i menneskelig hud forsynes af forskellige typer nervefibre.�Ann Neurol.�2005;58:88�95. doi: 10.1002/ana.20527.�[PubMed]�[Cross Ref]
35. Maricich SM, Morrison KM, Mathes EL, Brewer BM. Gnavere er afhængige af Merkel-celler til teksturdiskriminationsopgaver.�J Neurosci.�2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[PMC gratis artikel]�[PubMed]�[Cross Ref]
36. Ikeda I, Yamashita Y, Ono T, Ogawa H. Selektiv fototoksisk ødelæggelse af rotte Merkel-celler afskaffer reaktioner fra langsomt tilpassende type I-mekanoreceptorenheder.�J Physiol.�1994;479:247 56. [PMC gratis artikel][PubMed]
37. Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, et al. Merkel-celler er essentielle for lette berøringsresponser.�Videnskab.�2009;324:1580�2. doi: 10.1126/science.1172890.[PMC gratis artikel]�[PubMed]�[Cross Ref]
38. Diamond J, Holmes M, Sygeplejerske CA. Er Merkel celle-neurit gensidige synapser involveret i initieringen af taktile responser i salamanderhud?�J Physiol.�1986;376:101 20. [PMC gratis artikel][PubMed]
39. Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Spændingsafhængige strømme i isolerede enkelte Merkel-celler fra rotter.�J Physiol.�1992;450:143 62. [PMC gratis artikel]�[PubMed]
40. Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. Regelmæssigheden af vedvarende affyring afslører to populationer af langsomt tilpassende berøringsreceptorer i behåret hud på mus.�J Neurophysiol.�2010;103:3378�88. doi: 10.1152/jn.00810.2009.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
41. Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Immunlokalisering af aquaporin-1 i de mekanoreceptive Ruffini-ender i det parodontale ligament.�Hjerne res.�2007;1157:32�40. doi: 10.1016/j.brainres.2007.04.033.�[PubMed]�[Cross Ref]
42. Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K, et al. Påvisning af syrefølende ionkanal 3 (ASIC3) i parodontale Ruffini-ender på musefortænder.�Neurosci Lett.�2011;488:173�7. doi: 10.1016/j.neulet.2010.11.023.�[PubMed]�[Cross Ref]
43. Johnson KO. Roller og funktioner af kutane mekanoreceptorer.�Curr Opin Neurobiol.�2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed]�[Cross Ref]
44. Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A, et al. Transskriptionsfaktoren c-Maf styrer berøringsreceptorens udvikling og funktion.�Videnskab.�2012;335:1373�6. doi: 10.1126/science.1214314.�[PubMed]�[Cross Ref]
45. Mendelson M, Lowenstein WR. Mekanismer for receptortilpasning.�Videnskab.�1964;144:554�5. doi: 10.1126/science.144.3618.554.�[PubMed]�[Cross Ref]
46. Loewenstein WR, Mendelson M. Komponenter af receptortilpasning i en pacinian corpuscle.�J Physiol.�1965;177:377 97. [PMC gratis artikel]�[PubMed]
48. Basbaum AI, Jessell TM. Opfattelsen af smerte. I: Kandel ER, Schwartz JH, Jessell TM, red. Principper for neural videnskab. Fjerde udgave. McGraw-Hill compagies, 2000: 472-490.
49. Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A, et al. Mekanoreceptorundertyper med lav tærskel udtrykker selektivt MafA og er specificeret ved Ret-signalering.�Neuron.�2009;64:857�70. doi: 10.1016/j.neuron.2009.12.004.�[PubMed]�[Cross Ref]
50. Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. En rolle for Runx-transkriptionsfaktorsignalering i dorsale rodgangliesensoriske neurondiversifikation.�Neuron.�2006;49:379�93. doi: 10.1016/j.neuron.2006.01.008.�[PubMed]�[Cross Ref]
51. Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Molekylær identifikation af hurtigt tilpassende mekanoreceptorer og deres udviklingsmæssige afhængighed af ret signalering.�Neuron.�2009;64:841�56. doi: 10.1016/j.neuron.2009.11.003.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
52. Vallbo AB, Hagbarth KE. Aktivitet fra hudmekanoreceptorer registreret perkutant hos vågne mennesker.�Exp Neurol.�1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed]�[Cross Ref]
53. Macefield VG. Fysiologiske egenskaber ved lavtærskelmekanoreceptorer i led, muskler og hud hos mennesker.�Clin Exp Pharmacol Physiol.�2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed]�[Cross Ref]
54. Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Ca2+-bølger i keratinocytter overføres til sensoriske neuroner: involvering af ekstracellulær ATP- og P2Y2-receptoraktivering.�Biochem J.�2004;380:329�38. doi: 10.1042/BJ20031089.�[PMC gratis artikel]�[PubMed][Cross Ref]
55. Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. ATP-signalering er afgørende for humane keratinocytters respons på mekanisk stimulering ved hypo-osmotisk shock.�Exp Dermatol.�2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed]�[Cross Ref]
56. Amano M, Fukata Y, Kaibuchi K. Regulering og funktioner af Rho-associeret kinase.�Exp Cell Res.�2000;261:44�51. doi: 10.1006/excr.2000.5046.�[Anmeldelse]�[PubMed]�[Cross Ref]
57. Koyama T, Oike M, Ito Y. Inddragelse af Rho-kinase og tyrosinkinase i hypotonisk stress-induceret ATP-frigivelse i bovine aorta-endotelceller.�J Physiol.�2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
59. McCarter GC, Reichling DB, Levine JD. Mekanisk transduktion af rotte dorsale rodganglieneuroner in vitro.�Neurosci Lett.�1999;273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[PubMed]�[Cross Ref]
60. Drew LJ, Wood JN, Cesare P. Distinkte mekanosensitive egenskaber af capsaicinfølsomme og -ufølsomme sensoriske neuroner.�J Neurosci.�2002;22:RC228.�[PubMed]
61. Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P, et al. Syrefølende ionkanaler ASIC2 og ASIC3 bidrager ikke til mekanisk aktiverede strømme i pattedyrs sensoriske neuroner.�J Physiol.�2004;556:691�710. doi: 10.1113/jphysiol.2003.058693.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
62. McCarter GC, Levine JD. Ionisk basis for en mekanotransduktionsstrøm i voksne rotte dorsale rodganglieneuroner.�Mol Smerte.�2006;2:28. doi: 10.1186/1744-8069-2-28.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
63. Coste B, Crest M, Delmas P. Farmakologisk dissektion og fordeling af NaN/Nav1.9, T-type Ca2+ strømme og mekanisk aktiverede kationstrømme i forskellige populationer af DRG neuroner.�J Gen Physiol.�2007;129:57�77. doi: 10.1085/jgp.200609665.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
64. Hao J, Delmas P. Flere desensibiliseringsmekanismer af mekanotransducerkanaler former affyring af mekanosensoriske neuroner.�J Neurosci.�2010;30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[PubMed]�[Cross Ref]
65. Drew LJ, Wood JN. FM1-43 er en permeant blokering af mekanosensitive ionkanaler i sensoriske neuroner og hæmmer adfærdsreaktioner på mekaniske stimuli.�Mol Smerte.�2007;3:1. doi: 10.1186/1744-8069-3-1.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
66. Hao J, Delmas P. Registrering af mekanofølsomme strømme ved hjælp af piezoelektrisk drevet mekanostimulator.�Nat Protoc.�2011;6:979�90. doi: 10.1038/nprot.2011.343.�[PubMed]�[Cross Ref]
68. Hu J, Lewin GR. Mekanofølsomme strømme i neuritterne i dyrkede musesensoriske neuroner.�J Physiol.�2006;577:815�28. doi: 10.1113/jphysiol.2006.117648.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
69. Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. Radial strækning afslører distinkte populationer af mekanosensitive pattedyrs somatosensoriske neuroner.�Proc Natl Acad Sci US A. 2008;105:20015�20. doi: 10.1073/pnas.0810801105.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
70. Crawford AC, Evans MG, Fettiplace R. Aktivering og tilpasning af transducerstrømme i skildpaddes hårceller.�J Physiol.�1989;419:405 34. [PMC gratis artikel]�[PubMed]
71. Ricci AJ, Wu YC, Fettiplace R. Den endogene calciumbuffer og tidsforløbet for transducertilpasning i auditive hårceller.�J Neurosci.�1998;18:8261 77. [PubMed]
72. Vollrath MA, Kwan KY, Corey DP. Mikromaskineriet af mekanotransduktion i hårceller.�Annu Rev Neurosci.�2007;30:339�65. doi: 10.1146/annurev.neuro.29.051605.112917.�[Anmeldelse]�[PMC gratis artikel]�[PubMed]�[Cross Ref]
74. Waldmann R, Lazdunski MH. H(+)-gatede kationkanaler: neuronale syresensorer i NaC/DEG-familien af ionkanaler.�Curr Opin Neurobiol.�1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed]�[Cross Ref]
75. Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, et al. Ionkanalen ASIC1 bidrager til visceral, men ikke kutan mekanoreceptorfunktion.�Gastroenterologi.�2004;127:1739�47. doi: 10.1053/j.gastro.2004.08.061.�[PubMed]�[Cross Ref]
76. Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, et al. DRASIC-kationkanalen bidrager til påvisning af kutan berøring og syrestimuli hos mus.�Neuron.�2001;32:1071�83. doi: 10.1016/S0896-6273(01)00547-5.�[Erratum in: Neuron 2002 Jul 18;35] [2]�[PubMed]�[Cross Ref]
77. Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M, et al. Knockout af ASIC2-kanalen hos mus svækker ikke kutan mekanosensation, visceral mekanociception og hørelse.�J Physiol.�2004;558:659�69. doi: 10.1113/jphysiol.2004.066001.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
78. Damann N, Voets T, Nilius B. TRP'er i vores sanser.�Curr Biol.�2008;18:R880�9. doi: 10.1016/j.cub.2008.07.063.�[Anmeldelse]�[PubMed]�[Cross Ref]
79. Christensen AP, Corey DP. TRP-kanaler i mekanosensation: direkte eller indirekte aktivering?�Nat Rev Neurosci.�2007;8:510�21. doi: 10.1038/nrn2149.�[Anmeldelse]�[PubMed]�[Cross Ref]
80. Liedtke W, Tobin DM, Bargmann CI, Friedman JM. Mammalian TRPV4 (VR-OAC) styrer adfærdsmæssige reaktioner på osmotiske og mekaniske stimuli i Caenorhabditis elegans.�Proc Natl Acad Sci US A. 2003;100(Suppl 2):14531�6. doi: 10.1073/pnas.2235619100.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
81. Suzuki M, Mizuno A, Kodaira K, Imai M. Nedsat trykfornemmelse hos mus, der mangler TRPV4.�J Biol Chem. 2003;278:22664�8. doi: 10.1074/jbc.M302561200.�[PubMed]�[Cross Ref]
82. Liedtke W, Choe Y, Mart�-Renom MA, Bell AM, Denis CS, Sali A, et al. Vanilloid receptor-relateret osmotisk aktiveret kanal (VR-OAC), en kandidat hvirveldyr osmoreceptor.�Celle.�2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
83. Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Forbigående receptorpotentiale vanilloid 4 er essentiel ved kemoterapi-induceret neuropatisk smerte hos rotter.�J Neurosci.�2004;24:4444�52. doi: 10.1523/JNEUROSCI.0242-04.2004.�[Erratum i: J Neurosci. 2004 Jun;24] [23][PubMed]�[Cross Ref]
84. Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, et al. TRPA1 medierer de inflammatoriske virkninger af miljøirriterende stoffer og proalgetiske midler.�Celle.�2006;124:1269�82. doi: 10.1016/j.cell.2006.02.023.�[PubMed]�[Cross Ref]
85. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ, et al. TRPA1 bidrager til kulde, mekanisk og kemisk nociception, men er ikke afgørende for hårcelletransduktion.�Neuron.�2006;50:277�89. doi: 10.1016/j.neuron.2006.03.042.�[PubMed]�[Cross Ref]
86. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 og Piezo2 er væsentlige komponenter i særskilte mekanisk aktiverede kationkanaler.�Videnskab.�2010;330:55�60. doi: 10.1126/science.1193270.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
87. Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS, et al. Piezoproteiner er poredannende underenheder af mekanisk aktiverede kanaler.�Naturen.�2012;483:176�81. doi: 10.1038/nature10812.[PMC gratis artikel]�[PubMed]�[Cross Ref]
88. Bae C, Sachs F, Gottlieb PA. Den mekanosensitive ionkanal Piezo1 hæmmes af peptidet GsMTx4.�Biokemi.�2011;50:6295�300. doi: 10.1021/bi200770q.�[PMC gratis artikel]�[PubMed][Cross Ref]
89. Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. Drosophila Piezos rolle i mekanisk nociception.�Naturen.�2012;483:209�12. doi: 10.1038/nature10801.�[PMC gratis artikel]�[PubMed][Cross Ref]
90. Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. Mutationer i mekanotransduktionsproteinet PIEZO1 er forbundet med arvelig xerocytose.�Blod.�2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
91. Kawashima Y, G�l�oc GS, Kurima K, Labay V, Lelli A, Asai Y, et al. Mekanotransduktion i musens indre øre-hårceller kræver transmembrankanallignende gener.�J Clin Invest.�2011;121: 4796�809. doi: 10.1172/JCI60405.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
92. Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouch�ne J, et al. TMC1, men ikke TMC2, er ansvarlig for autosomal recessiv ikke-syndromisk hørenedsættelse i tunesiske familier.�Audiol Neurootol.�2008;13:213�8. doi: 10.1159/000115430.�[PubMed]�[Cross Ref]
93. Manji SS, Miller KA, Williams LH, Dahl HH. Identifikation af tre nye høretab musestammer med mutationer i Tmc1-genet.�Am J Pathol.�2012;180:1560�9. doi: 10.1016/j.ajpath.2011.12.034.[PubMed]�[Cross Ref]
94. Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, et al. Et stomatin-domæneprotein, der er afgørende for berøringsfornemmelse i musen.�Naturen.�2007;445:206�9. doi: 10.1038/nature05394.�[PubMed][Cross Ref]
95. Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, et al. Stomatin og sensorisk neuronmekanotransduktion.�J Neurophysiol.�2007;98:3802�8. doi: 10.1152/jn.00860.2007.[PubMed]�[Cross Ref]
96. Huang M, Gu G, Ferguson EL, Chalfie M. Et stomatin-lignende protein, der er nødvendigt for mekanosensation hos C. elegans.�Naturen.�1995;378:292�5. doi: 10.1038/378292a0.�[PubMed]�[Cross Ref]
97. Hu J, Chiang LY, Koch M, Lewin GR. Bevis for en proteinbinding involveret i somatisk berøring.�EMBO J.�2010;29:855�67. doi: 10.1038/emboj.2009.398.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
98. Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L, et al. Laminin-332 koordinerer mekanotransduktion og vækstkeglebifurkation i sensoriske neuroner.�Nat Neurosci.�2011;14: 993�1000. doi: 10.1038/nn.2873.�[PubMed]�[Cross Ref]
99. Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, et al. TWIK-1, en allestedsnærværende menneskelig, svagt indadrettet K+-kanal med en ny struktur.�EMBO J.�1996;15:1004 11.[PMC gratis artikel]�[PubMed]
100. Lesage F. Farmakologi af neuronale baggrundkaliumkanaler.�Neurofarmakologi.�2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed]�[Cross Ref]
101. Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE, et al. Fordelingsanalyse af humane kaliumkanaler i to poredomæner i væv i centralnervesystemet og periferi.�Brain Res Mol Brain Res.�2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Cross Ref]
102. Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor� E. Mekano- eller syrestimulering, to interaktive aktiveringsmåder af TREK-1 kaliumkanalen.�J Biol Chem. 1999;274:26691�6. doi: 10.1074/jbc.274.38.26691.�[PubMed]�[Cross Ref]
103. Maingret F, Fosset M, Lesage F, Lazdunski M, Honor� E. TRAAK er en pattedyrs neuronal mekano-gated K+-kanal.�J Biol Chem. 1999;274:1381�7. doi: 10.1074/jbc.274.3.1381.�[PubMed][Cross Ref]
104. Alloui A, Zimmermann K, Mamet J, Duprat F, No�l J, Chemin J, et al. TREK-1, en K+-kanal involveret i polymodal smerteopfattelse.�EMBO J.�2006;25:2368�76. doi: 10.1038/sj.emboj.7601116.[PMC gratis artikel]�[PubMed]�[Cross Ref]
105. No�l J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, et al. De mekano-aktiverede K+-kanaler TRAAK og TREK-1 styrer både varm og kold opfattelse.�EMBO J.�2009;28:1308�18. doi: 10.1038/emboj.2009.57.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
106. Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. TRESK to-pore-domæne K+ kanaler udgør en væsentlig komponent i baggrunden af kaliumstrømme i murine dorsale rodganglieneuroner.�J Physiol.�2007;585:867�79. doi: 10.1113/jphysiol.2007.145649.[PMC gratis artikel]�[PubMed]�[Cross Ref]
107. Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR, et al. Stigende midler fra Szechuan peberfrugter ophidser sensoriske neuroner ved at hæmme to-pore kaliumkanaler.�Nat Neurosci.�2008;11: 772�9. doi: 10.1038/nn.2143.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
108. Lennertz RC, Tsunozaki M, Bautista DM, Stucky CL. Fysiologisk grundlag for prikkende paræstesi fremkaldt af hydroxy-alpha-sanshool.�J Neurosci.�2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[PMC gratis artikel]�[PubMed]�[Cross Ref]
109. Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM, et al. KCNQ4 K(+) kanaler tuner mekanoreceptorer til normal berøringsfornemmelse hos mus og mennesker.�Nat Neurosci.�2012;15: 138�45. doi: 10.1038/nn.2985.�[PubMed]�[Cross Ref]
110. Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG, et al. Et genetisk grundlag for mekanosensoriske træk hos mennesker.�PLoS Biol.�2012;10:e1001318. doi: 10.1371/journal.pbio.1001318.[PMC gratis artikel]�[PubMed]�[Cross Ref]
111. Delmas P, Hao J, Rodat-Despoix L. Molekulære mekanismer for mekanotransduktion i pattedyrs sensoriske neuroner.�Nat Rev Neurosci.�2011;12:139�53. doi: 10.1038/nrn2993.�[PubMed]�[Cross Ref]
Mekanoreceptive smerter: Ifølge CDC havde �mere end 50 % af amerikanske voksne (125 millioner) et muskuloskeletal smertelidelse i 2012.�
�Mere end 40 % af voksne med en muskel- og knoglelidelse brugte en komplementær sundhedstilgang af en eller anden grund i 2012. Dette var signifikant højere end brugen blandt personer uden en muskel- og knoglesmerter (24.1 %). Brugen af komplementære sundhedsmetoder af en eller anden grund blandt personer med nakkesmerter eller problemer var mere end dobbelt så høj som brugen blandt personer uden disse problemer.�
�Blandt voksne med en lidelse i bevægeapparatet var brugen af en komplementær sundhedstilgang højest blandt dem med nakkesmerter eller problemer (50.6 %), efterfulgt af personer med andre muskel- og knogleproblemer (46.2 %).
�Brugen af komplementære sundhedstilgange af en eller anden grund blandt personer med nakkesmerter eller problemer var mere end dobbelt så høj som brugen blandt personer uden disse problemer.�
Mekanoreceptorer er sensoriske receptorer, der reagerer på mekanisk tryk eller forvrængning.
Disse omfatter kutane receptorer til berøring, receptorer, der overvåger muskellængde og spændinger, auditive og vestibulære receptorer og andre.
GATE KONTROL TEORI OM SMERTE
Ikke-smertefuldt input lukker portene for smertefuldt input.
Dette forhindrer smertefornemmelser i at rejse til højere kortikale niveauer
Afferenter med lille diameter (smerte) hæmmer hæmning af smerte
Afferenter med stor diameter (vibration) har en tendens til at fremkalde hæmning af smerte.
Denne teori hævder, at ikke-nociceptive fibre kan forstyrre signaler fra smertefibre og derfor hæmme smerte.
Stor diameter A? fibre er ikke-nociceptive (transmitterer ikke smertestimuli) og hæmmer virkningerne af affyring af A? og C-fibre.
DORSALSØLE MEDIAL LEMNISKAL VEJ
UDNYTTELSE AF PERIFERE MEKANORECEPTORER TIL AT ÆNDRE SMERTEPERCEPTION
Hvordan kan vi hjælpe?
TILBAGETRÆKNINGSREFLEKS
En exciteret afferent neuron stimulerer excitatoriske interneuroner, som igen stimulerer de efferente motoriske neuroner, der forsyner biceps, den muskel i armen, der bøjer (bøjer) albueleddet. Sammentrækning af biceps trækker hånden væk fra den varme komfur.
Den afferente neuron stimulerer også hæmmende interneuroner, som igen hæmmer de efferente neuroner, der forsyner triceps for at forhindre, at den trækker sig sammen. Denne type neuronal forbindelse, der involverer stimulering af nerveforsyningen til en muskel og samtidig hæmning af nerverne til dens antagonistiske muskel, er kendt som gensidig hæmning.
Den afferente neuron stimulerer stadig andre interneuroner, der fører signalet op af rygmarven til hjernen via en stigende vej. Først når impulsen når det sensoriske område af cortex, er personen opmærksom på smerten, dens placering og typen af stimulus. Desuden, når impulsen når hjernen, kan informationen lagres som hukommelse, og personen kan tænke over, hvad der skete.
RECEPTORBASERET TERAPI
Reguleringer
Aktivering af ledmekanoreceptorer gennem kiropraktiske justeringer kan modulere og �overskygge� hjernens opfattelse af fibre med mindre diameter.
Gentagelse af aktivering af ledmekanoreceptorer kan skabe positiv plasticitet i afferente veje.
Positiv plasticitet kan lukke ned for smerte
Vibration
Vibrationsstimulering ved specifikke frekvenser kan ændre smerteopfattelsen
Gentagelse af aktivering af Merkels diske og Meissners blodlegemer kan skabe positiv plasticitet i afferente veje.
Igen kan positiv plasticitet lukke smerten ned
VIBRATION
�Denne type enhed anvender sinusformede vibrationer og tilbyder kontinuerlig valgbar amplitude på 0-5.2 mm afhængigt af fodpositionen og en valgbar frekvens på 5-30Hz.�
�WBV-træning ser ud til at være en effektiv, sikker og passende intervention for siddende arbejdende medarbejdere med kroniske lændesmerter.�
� Homotopisk vibro-taktil stimulering resulterede i 40 % varmesmertereduktion i alle emnegrupper. Distraktion så ikke ud til at påvirke eksperimentelle smertevurderinger.�
�Vibro-taktil stimulering rekrutterede effektivt analgetiske mekanismer ikke kun i NC, men også hos patienter med kroniske muskuloskeletale smerter, herunder FM.�
LET BØR
�I alt oplevede 44 raske frivillige varmesmerter og CT-optimale (langsom børstning) og CT-suboptimale (hurtig børstning eller vibration) stimuli. Tre forskellige eksperimentelle paradigmer blev brugt: Samtidig påføring af varmesmerter og taktil (langsom børstning eller vibration) stimulering; Langsom børstning, påført i variabel varighed og intervaller, forud for varmesmerter; Langsom kontra hurtig børstning forud for varmesmerter.�
Hos mennesker tilhører de vigtigste hjerneområder, der modtager C-LTMR-information, det somatosensoriske system og påvirker behandlende hjernenetværk som den kontralaterale bageste insulære cortex eller den mediale præfrontale cortex. Intensiteten af CT-målrettet berøring er kodet i den primære og sekundære somatosensoriske cortex (S1 kontralateral, S2 bilateral), hvorimod behageligheden er kodet i den forreste cingulate cortex. C-LTMR'er aktiverer også regioner involveret i belønningsbehandling (putamen og orbitofrontal cortex) og i bearbejdning af sociale stimuli (posterior superior temporal sulcus).
ALT PERIFERT HAR EN CENTRAL KONSEKVENS
CASESTUDIE
47-årig mand fik venstre CVA i oktober 2017.
Har ikke bevæget højre side af kroppen siden ulykken.
Præsenteret til vores klinik, fordi han ønsker at �vende tilbage til det.�
FYSISK UNDERSØGELSE HØJDEPUNKTER
Dysartri
Ændret smerteopfattelse
Svært ved simpel matematik
Slap på RUE og RLE
FYSISK UNDERSØGELSE HØJDEPUNKTER
Patienten havde ingen bevægelse, før vi begyndte at teste fornemmelse og reflekser:
ALLODYNI:�Hviser til central smertesensibilisering (øget reaktion fra neuroner) efter normalt ikke-smertefuld, ofte gentagne stimulering.
Allodyni kan føre til udløsning af en smerterespons fra stimuli, som normalt ikke fremkalder smerte.
Temperatur eller fysisk stimuli kan fremkalde allodyni, som kan føles som en brændende fornemmelse, og det opstår ofte efter skade på et sted.
Alldynia er forskellig fra Hyperalgesi, en ekstrem, overdreven reaktion på en stimulus, som normalt er smertefuld.
Hvorfor fører lokal skade eller skade forårsaget af traumer til kroniske, vanskelige smerter hos visse patienter? Hvad er ansvarlig for oversættelsen af lokal skade med akutte smerter til en kronisk smertetilstand? Hvorfor reagerer nogle smerter på antiinflammatoriske lægemidler og/eller medicin, mens andre former for smerte kræver opiater?
Smerte er en indviklet proces, der involverer både det perifere nervesystem (PNS) og centralnervesystemet (CNS). Vævsskade udløser PNS, som transmitterer signaler via rygmarven ind i hjernen, hvor smerteopfattelsen opstår. Men hvad får den intense smerteoplevelse til at udvikle sig til et uophørligt fænomen? Kan der gøres noget for at forhindre det? Beviser tyder på det kronisk smerte resultater fra en kombination af mekanismer, såsom neurologiske "minder" om forudgående smerte.
Nociception: Den enkleste vej
Akutte eller nociceptive smerter er karakteriseret som den regelmæssige oplevelse af ubehag, der opstår som reaktion på meget grundlæggende skade eller skade. Det er beskyttende og advarer os om at bevæge os væk fra fornærmelsens oprindelse og tage os af traumet. De mekanismer, der skaber nociceptiv smerte omfatter transduktion, som udvider den eksterne traumatiske stimulering til elektrisk aktivitet i specialiserede nociceptive primære afferente nerver. De afferente nerver leder derefter den sensoriske information fra PNS til CNS.
I CNS transmitteres smertedataene af de primære sensoriske neuroner til centrale projektionsceller. Efter at informationen er overført til alle de områder af hjernen, som er ansvarlige for vores perception, sker den faktiske sanseoplevelse. Nociceptive smerter er en relativt simpel reaktion på en særlig simpel, akut stimulus. Men mekanikken, der er ansvarlig for nociceptiv smerte, kan ikke identificere fænomener, såsom smerte, der fortsætter på trods af fjernelse eller heling af stimulationen, såsom i tilfælde af fantomsmerter i lemmer.
Smerter og den inflammatoriske reaktion
I tilfælde af mere alvorlige skader, såsom operationssår, kan vævsskade stimulere en inflammatorisk reaktion. Men andre tilstande, især gigt, kan også karakteriseres ved vedvarende tilfælde af betændelse forbundet med intense smertesymptomer. Mekanismerne for denne type smerte relateret til vævsbeskadigelse og en inflammatorisk respons er forskellige fra nociceptive smerter med tidlig advarsel.
Når man observerer snittet eller stedet for anden skade eller skade, opstår der en kaskade af hyperexcitable hændelser i nervesystemet. Dette kropslige "wind-up"-fænomen begynder ved huden, hvor det forstærkes langs de perifere nerver, og kulminerer med en overfølsomhedsreaktion langs rygmarven (dorsalhornet) og hjernen. Inflammatoriske celler omgiver derefter områderne med vævsskade og producerer også cytokiner og kemokiner, stoffer, der er beregnet til at mediere helingsprocessen og vævsregenerering. Men disse midler kan også betragtes som irriterende og justere egenskaberne af de primære sensoriske neuroner, der omgiver traumeområdet.
De væsentligste faktorer, som udløser inflammatorisk smerte, omfatter således beskadigelse af nociceptorerne med høj tærskel, kendt som perifer sensibilisering, ændringer og ændringer af neuronerne i nervesystemet og forstærkningen af excitabiliteten af neuroner i CNS. Dette repræsenterer central sensibilisering og er ansvarlig for overfølsomhed, hvor områder, der støder op til dem af den sande skade, vil opleve smerte, som om disse var skadet. Disse væv kan også reagere på stimulering, som normalt ikke skaber smerte, såsom en berøring, iført tøj, let tryk eller endda børstning af dit eget hår, som om de virkelig var smertefulde, kaldet allodyni.
Neuropatisk smerte skyldes beskadigelse eller skade på nervesystemet, såsom karpaltunnelsyndrom, postherpetisk neuralgi og diabetisk neuropati. Selvom nogle af de mekanismer, der synes at forårsage neuropatisk smerte, overlapper med dem, der er ansvarlige for inflammatorisk smerte, er mange af dem forskellige og vil derfor have brug for en anden tilgang til deres behandling.
Processen med perifer og central sensibilisering opretholdes, i det mindste teoretisk og eksperimentelt, under den excitatoriske neurotransmitter, glutamat, som menes at blive frigivet, når N-methyl-D-aspartat (NMDA)-receptoren aktiveres.
Nervesystemet består af enten hæmmende eller excitatoriske neurotransmittere. Det meste af det, der tillader vores nervesystem at reagere passende på skader eller skader, er finjustering eller hæmning af en række processer. Overexcitation af nervesystemet ses at være et problem i en række forskellige lidelser. For eksempel kan overaktivering af en NMDA-receptor også være relateret til affektive lidelser, sympatiske abnormiteter og endda opiattolerance.
Selv almindelig nociceptiv smerte aktiverer til en vis grad NMDA-receptoren og menes at føre til glutamatfrigivelse. Ikke desto mindre er overfølsomhed over for NMDA-receptoren nøglen til neuropatisk smerte.
Med andre typer kroniske smerter, såsom fibromyalgi og spændingshovedpine, kan nogle af de mekanismer, der er aktive ved inflammatoriske og neuropatiske smerter, også skabe lignende abnormiteter i smertesystemet, herunder central sensibilisering, højere excitabilitet af de somatosensoriske veje og reduktioner i hæmmende mekanismer i centralnervesystemet.
Perifer sensibilisering
Cyclo-oxygenase (COX) spiller også en væsentlig funktion i både perifere og centrale sensibiliseringer. COX-2 er et af de enzymer, som induceres under den inflammatoriske proces; COX-2 omdanner arachidonsyre til prostaglandiner, hvilket øger følsomheden af perifere nociceptorterminaler. Stort set forårsager perifer inflammation også, at COX-2 produceres fra CNS. Signaler fra perifere nociceptorer er delvist ansvarlige for denne opregulering, men der synes også at være en humoral komponent i transduktionen af smertesignalerne over blod-hjerne-barrieren.
For eksempel, i eksperimentelle modeller, genereres COX-2 fra CNS, selvom dyr modtager en sensorisk nerveblokering før perifer inflammatorisk stimulering. COX-2, der udtrykkes over de dorsale hornneuroner i rygmarven, frigiver prostaglandiner, som virker på de centrale terminaler eller de præsynaptiske terminaler af nociceptive sensoriske fibre for at øge transmitterfrigivelsen. Derudover virker de postsynaptisk på de dorsale hornneuroner for at producere direkte depolarisering. Og endelig hæmmer de aktiviteten af glycinreceptoren, og dette er en hæmmende transmitter. Derfor skaber prostaglandinerne en stigning i excitabilitet af centrale neuroner.
Hjerneplasticitet og central sensibilisering
Central sensibilisering beskriver ændringer, der sker i hjernen som reaktion på gentagen nervestimulation. Efter gentagne stimuli ændres mængden af hormoner og hjernens elektriske signaler, efterhånden som neuroner udvikler en "hukommelse" til at reagere på disse tegn. Konstant stimulering skaber en mere kraftfuld hjernehukommelse, så hjernen vil reagere hurtigere og mere effektivt, når den i fremtiden gennemgår den samme stimulering. De deraf følgende ændringer i hjernens ledninger og reaktion omtales som neural plasticitet, som beskriver hjernens evne til let at ændre sig selv, eller central sensibilisering. Derfor aktiveres eller sensibiliseres hjernen af tidligere eller gentagne stimuli for at blive mere exciterbar.
Udsvingene af central sensibilisering opstår efter gentagne møder med smerte. Forskning i dyr indikerer, at gentagen udsættelse for en smertefuld stimulation vil ændre dyrets smertetærskel og føre til en stærkere smertereaktion. Forskere mener, at disse ændringer kan forklare den vedvarende smerte, der kan opstå selv efter en vellykket rygoperation. Selvom en diskusprolaps kan fjernes fra en klemt nerve, kan smerte fortsætte som et minde om nervekompressionen. Nyfødte, der gennemgår omskæring uden bedøvelse, vil reagere mere dybtgående på fremtidig smertefuld stimulering, såsom rutinemæssige injektioner, vaccinationer og andre smertefulde processer. Disse børn har ikke kun en højere hæmodynamisk reaktion, kendt som takykardi og takypnø, men de vil også udvikle øget gråd.
Denne neurologiske hukommelse af smerte blev grundigt undersøgt. I en rapport om sine tidligere forskningsundersøgelser bemærkede Woolf, at den forbedrede refleksexcitabilitet efter perifer vævsskade eller -skade ikke er afhængig af fortsatte perifere inputsignaler; i stedet for, timer efter et perifert traume, fortsatte de modtagelige felter af rygmarvshornneuroner med at forstørre. Forskere har også dokumenteret betydningen af den spinale NMDA-receptor for induktion og vedligeholdelse af central sensibilisering.
Betydning for smertebehandling
Når først central sensibilisering er etableret, kræves der ofte større doser af analgetika for at undertrykke den. Forebyggende analgesi eller terapi før smerten skrider frem, kan sænke virkningerne af alle disse stimuleringer på CNS. Woolf påviste, at den morfindosis, der kræves for at stoppe central hyperexcitabilitet, givet før kortvarig skadelig elektrisk stimulering hos rotter, var en tiendedel af den dosis, der krævedes for at afskaffe aktivitet, efter at den var vokset. Dette oversættes til klinisk praksis.
I et klinisk forsøg med 60 patienter, der gennemgår abdominal hysterektomi, krævede personer, der fik 10 mg morfin intravenøst på tidspunktet for induktion af anæstesi, signifikant mindre morfin til postoperativ smertekontrol. Ydermere var smertefølsomheden omkring såret, omtalt som sekundær hyperalgesi, også reduceret i den forbehandlede morfingruppe. Forebyggende analgesi blev brugt med sammenlignelig succes i en række kirurgiske indstillinger, herunder præspinal operation og postortopædisk operation.
En enkelt dosis på 40 eller 60 mg/kg rektal acetaminophen har en klar morfinbesparende effekt ved dagkirurgi hos børn, hvis det administreres i induktion af anæstesi. Desuden oplevede børn med tilstrækkelig analgesi med acetaminophen signifikant mindre postoperativ kvalme og opkastning.
NMDA-receptorantagonister har givet postoperativ analgesi, når de administreres præoperativt. Der findes forskellige rapporter i litteraturen, der understøtter brugen af ketamin og dextromethorphan i den præoperative periode. Hos patienter, der gennemgår forreste korsbåndsrekonstruktion, var 24-timers patientkontrolleret analgesiopioidforbrug signifikant mindre i den præoperative dextromethorphan-kategori versus placebogruppen.
I dobbeltblindede, placebokontrollerede forskningsstudier blev gabapentin indiceret som et præmedicinsk analgetikum til patienter, der gennemgår mastektomi og hysterektomi. Præoperativ oral gabapentin reducerede smertescore og postoperativt analgetikaforbrug uden forskel i bivirkninger sammenlignet med placebo.
Præoperativ administration af nonsteroide antiinflammatoriske lægemidler (NSAID'er) har vist et signifikant fald i opioidbrug postoperativt. COX-2'er er at foretrække på grund af deres relative mangel på blodpladeeffekter og betydelige gastrointestinale sikkerhedsprofiler sammenlignet med konventionelle NSAID'er. Celecoxib, rofecoxib, valdecoxib og parecoxib, uden for USA, administreret præoperativt reducerer postoperativ narkotikabrug med mere end 40 procent, hvor mange patienter bruger mindre end halvdelen af opioiderne sammenlignet med placebo.
Blokering af nerveledning i den præoperative periode ser ud til at forhindre udviklingen af central sensibilisering. Phantom limb syndrome (PLS) er blevet tilskrevet et spinal wind-up fænomen.�Patienter med amputation
har ofte brændende eller prikkende smerter i kropsdelen fjernet. En mulig årsag er, at nervetråde ved stumpen stimuleres, og hjernen fortolker signalerne som at de stammer fra den amputerede del. Den anden er omlejringen inden for de kortikale områder, så området siger for hånden nu reagerer på signaler fra andre dele af kroppen, men stadig tolker dem som at komme til den amputerede hånd.
For patienter, der blev amputeret i den nedre ekstremitet under epidural anæstesi, udviklede ingen af de 11 patienter, der fik lumbal epidural blokade med bupivacain og morfin i 72 timer før operationen imidlertid PLS. For personer, der gennemgik generel anæstesi uden forudgående lumbal epidural blokade, havde 5 ud af 14 patienter PLS efter 6 uger og 3 fortsatte med at opleve PLS efter 1 år.
Woolf og Chong har bemærket, at perfekt præoperativ, intraoperativ og postoperativ behandling består af "NSAID'er for at reducere aktiveringen/centraliseringen af nociceptorer, lokalbedøvelse til at blokere sensorisk indstrømning og centralt virkende lægemidler såsom opiater." Formindskelse af perioperativ smerte med forebyggende teknikker øger tilfredsheden, fremskynder udflåd, skåner opioidbrug, sammen med mindsket forstoppelse, sedation, kvalme og urinretention og kan endda stoppe udviklingen af kronisk smerte. Anæstesilæger og kirurger bør overveje at integrere disse teknikker i deres daglige praksis.
Når smerte opstår som følge af beskadigelse eller skade som følge af operation, kan rygmarven opnå en hyperexciterbar tilstand, hvor der opstår overdrevne smertereaktioner, som kan vare ved i dage, uger eller endda år.
Hvorfor resulterer lokaliseret skade som følge af traumer i kroniske, vanskelige smerter hos nogle patienter? Vævsskade fører til en konstellation af ændringer i spinal excitabilitet, herunder forhøjet spontan affyring, større responsamplitude og -længde, nedsat tærskel, øget udledning til gentagen stimulation og udvidede modtagelige felter. Vedvaren af disse ændringer, som under ét kaldes central sensibilisering, ser ud til at være fundamental for den langvarige forbedring af smertefølsomheden, som definerer kronisk smerte. Talrige lægemidler og/eller medicin samt lokalbedøvende neural blokade kan begrænse omfanget af det centrale nervesystem (CNS) vindup, hvilket fremgår af mindsket smerte og nedsat opioidforbrug i de forebyggende analgetiske modeller.
Dr. Alex Jimenez's Insight
Kiropraktisk pleje er en alternativ behandlingsmulighed, som bruger rygmarvsjusteringer og manuelle manipulationer til sikkert og effektivt at genoprette samt opretholde den korrekte justering af rygsøjlen. Forskningsundersøgelser har fastslået, at rygmarvsfejlstillinger eller subluksationer kan føre til kroniske smerter. Kiropraktisk behandling er almindeligvis brugt til smertebehandling, selvom symptomerne ikke er forbundet med en skade og/eller tilstand i bevægeapparatet og nervesystemet. Ved omhyggeligt at justere rygsøjlen igen, a kiropraktor kan hjælpe med at reducere stress og pres fra strukturerne omkring hovedkomponenten af kroppens fundament, hvilket i sidste ende giver smertelindring.
Det enteriske nervesystems funktion og smerter
Når det kommer til den reducerede brug af lægemidler og/eller medicin, herunder opioider, for at forhindre bivirkninger som gastrointestinale sundhedsproblemer, kan det enteriske nervesystems korrekte funktion være i spil.
Det enteriske nervesystem (ENS) eller det indre nervesystem er en af nøglegrenene af det autonome nervesystem (ANS) og består af et mesh-lignende system af nerver, som modulerer mave-tarmkanalens rolle. Det er i stand til at virke uafhængigt af det sympatiske og parasympatiske nervesystem, selvom det kan være påvirket af dem. ENS kan også kaldes den anden hjerne.�Den er afledt af neurale kamceller.
Det enteriske nervesystem hos mennesker består af omkring 500 millioner neuroner, inklusive de mange typer Dogiel-celler, cirka en to hundrededel af mængden af neuroner i hjernen. Det enteriske nervesystem indsættes i slimhinden i mave-tarmsystemet, begyndende ved spiserøret og strækker sig ned til anus. Dogiel-celler, også kendt som Dogiel-celler, refererer til en slags multipolært binyrevæv i de prævertebrale sympatiske ganglier.
ENS er i stand til at udføre autonome funktioner, såsom koordinering af reflekser; selvom det modtager betydelig innervation i det autonome nervesystem, gør det og kan fungere uafhængigt af hjernen og rygmarven.�Det enteriske nervesystem er blevet beskrevet som "den anden hjerne" af en række årsager. Det enteriske nervesystem kan fungere autonomt. Det kommunikerer normalt med centralnervesystemet (CNS) via det parasympatiske eller via vagusnerven og det sympatiske, det vil sige gennem de prævertebrale ganglier, nervesystemerne. Imidlertid afslører undersøgelser af hvirveldyr, at når vagusnerven afskæres, fortsætter det enteriske nervesystem med at fungere.
Hos hvirveldyr omfatter det enteriske nervesystem efferente neuroner, afferente neuroner og interneuroner, som alle gør det enteriske nervesystem i stand til at bære reflekser og fungere som et integrerende center i fravær af CNS-input. De sensoriske neuroner rapporterer om mekaniske og kemiske forhold. Det enteriske nervesystem har evnen til at ændre sin reaktion baseret på faktorer som næringsstof og bulksammensætning. Derudover indeholder ENS støtteceller, der ligner hjernens astroglia og en diffusionsbarriere omkring kapillærerne omkring ganglier, der ligner blod-hjerne-barrieren i blodkar.
Det enteriske nervesystem (ENS) spiller en central rolle i inflammatoriske og nociceptive processer. Lægemidler og/eller medicin, der interagerer med ENS, har for nylig vakt betydelig interesse på grund af deres evne til at regulere adskillige aspekter af tarmfysiologien og patofysiologien. Især dyreforsøg har vist, at proteinase-aktiverede receptorer (PAR'er) kan være afgørende for neurogen inflammation i tarmen. Desuden synes PAR2-agonister at inducere intestinal hypersensitivitet og hyperalgetiske tilstande, hvilket tyder på en rolle for denne receptor i visceral smerteopfattelse.
Desuden repræsenterer PAR'er sammen med proteinaserne, der aktiverer dem, spændende nye mål for terapeutisk intervention på ENS. Omfanget af vores information er begrænset til kiropraktik samt til rygmarvsskader og -tilstande. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emner: Ischias
Iskias er medicinsk omtalt som en samling af symptomer, snarere end en enkelt skade og / eller tilstand. Symptomer på sciatic nervesmerter eller ischias kan variere i hyppighed og intensitet, men det er oftest beskrevet som en pludselig, skarp (knivlignende) eller elektrisk smerte, der udstråler fra den lave ryg ned mellem balder, hofter, lår og lår benene i foden. Andre symptomer på iskias kan omfatte, prikkende eller brændende fornemmelser, følelsesløshed og svaghed langs længden af den sciatic nerve. Ischias påvirker oftest individer i alderen af 30 og 50 år. Det kan ofte udvikle sig som følge af degenerationen af rygsøjlen på grund af alder, men komprimeringen og irritationen af den sciatic nerve forårsaget af en bulging eller herniated disc, blandt andet spinal sundhed problemer, kan også forårsage sciatic nerve smerte.
IFM's Find A Practitioner-værktøj er det største henvisningsnetværk inden for Functional Medicine, skabt for at hjælpe patienter med at finde Functional Medicine-praktikere overalt i verden. IFM Certified Practitioners er anført først i søgeresultaterne på grund af deres omfattende uddannelse i Functional Medicine

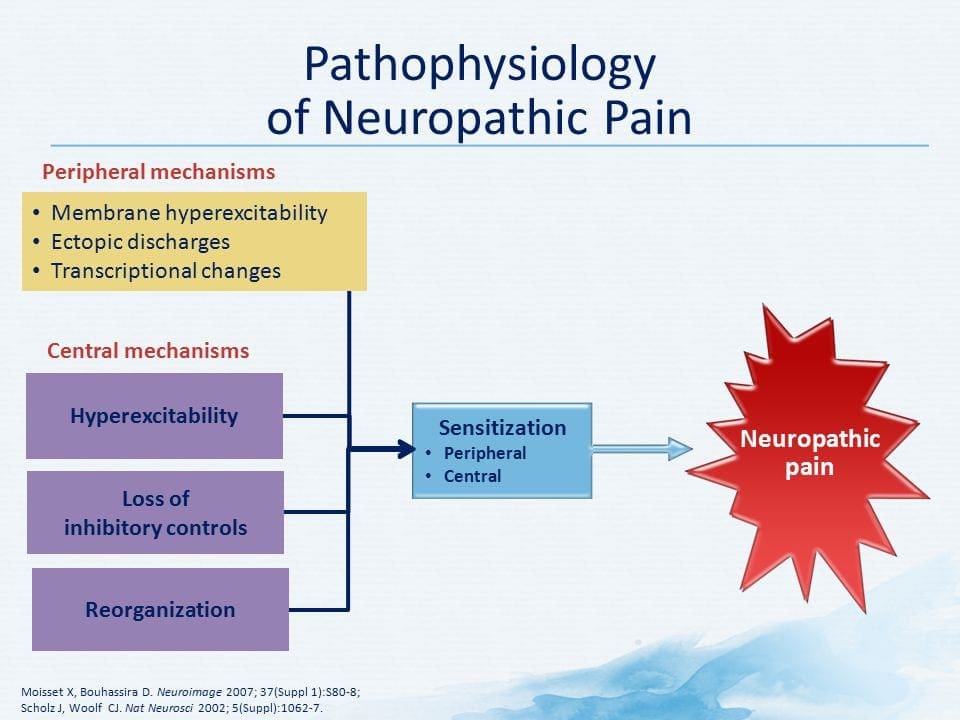
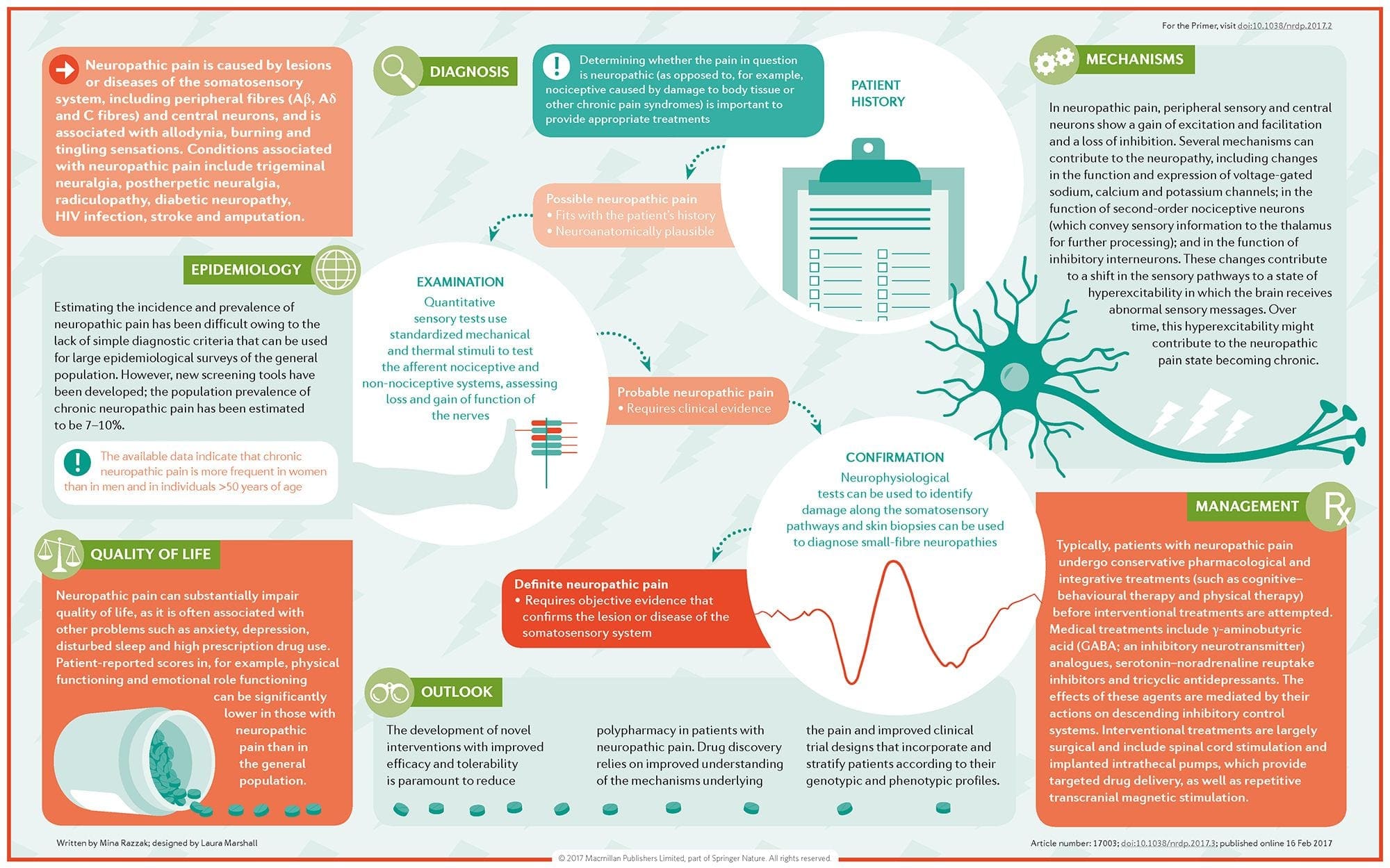




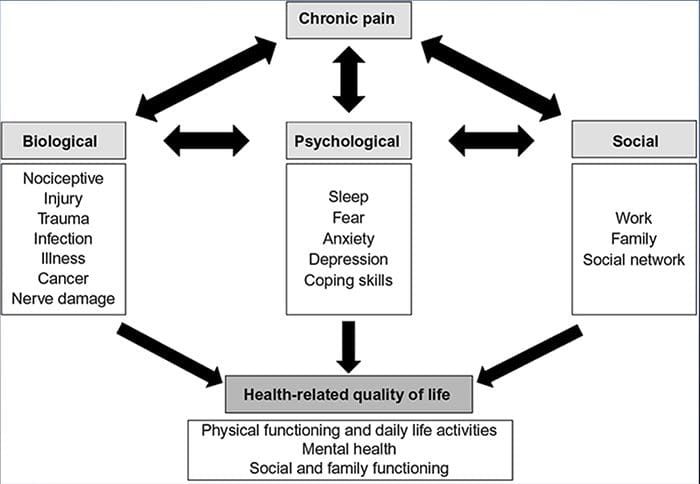
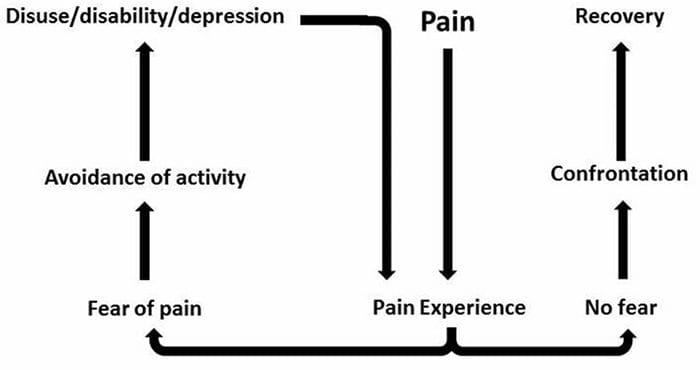
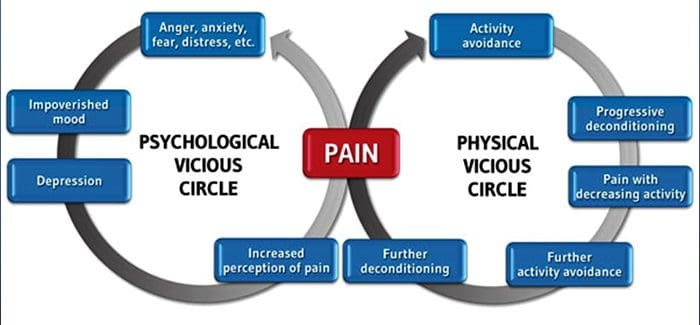
















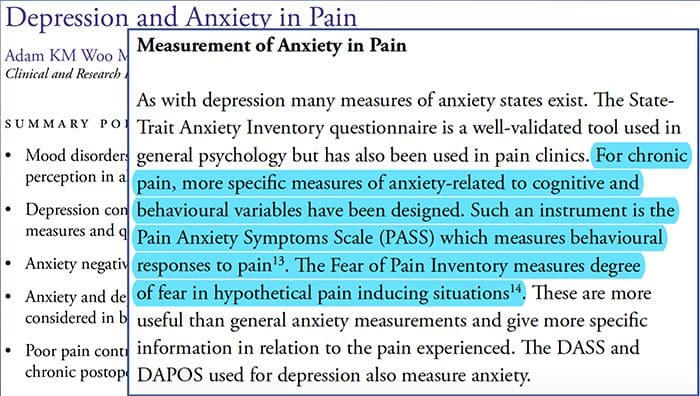


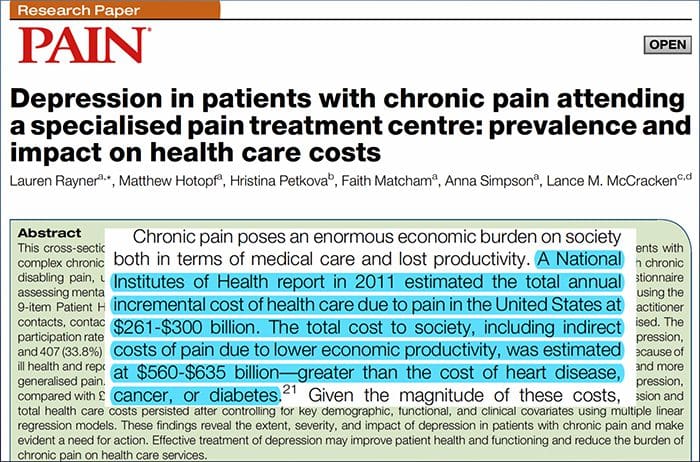



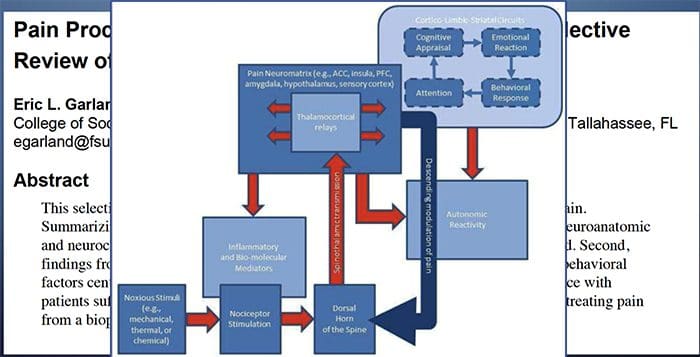






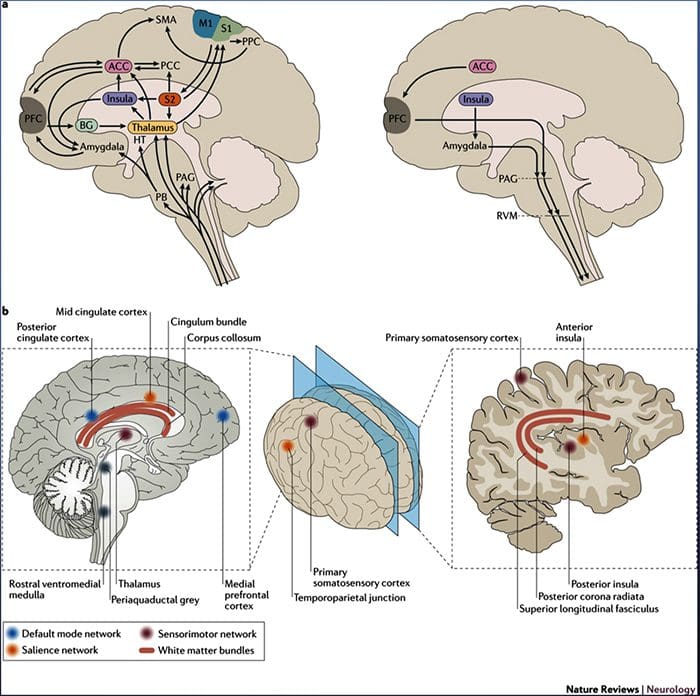

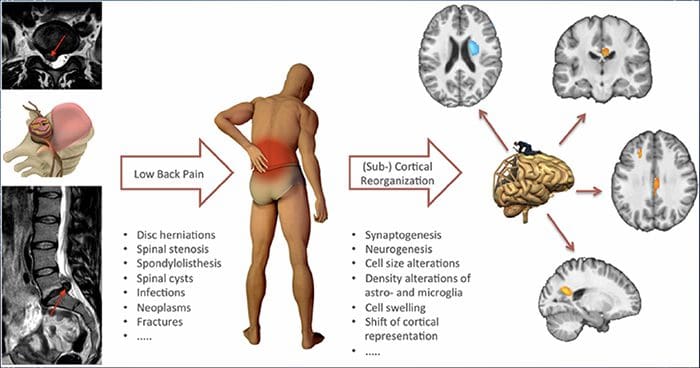

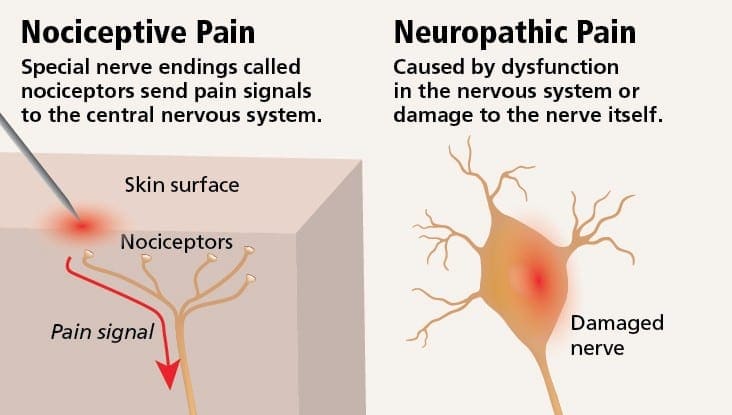
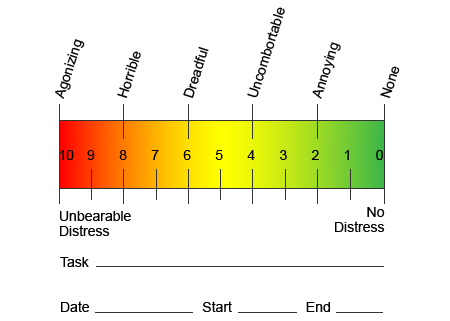


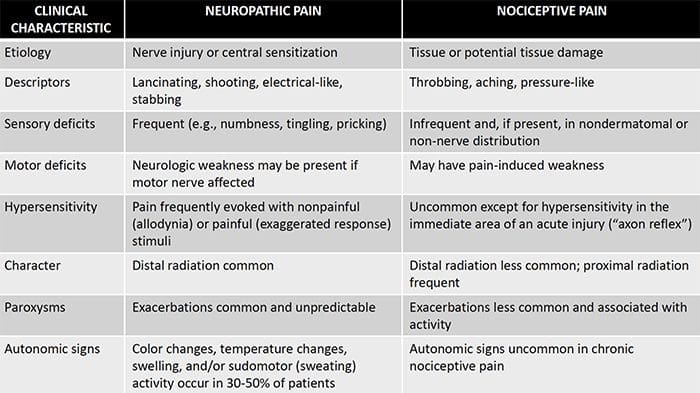
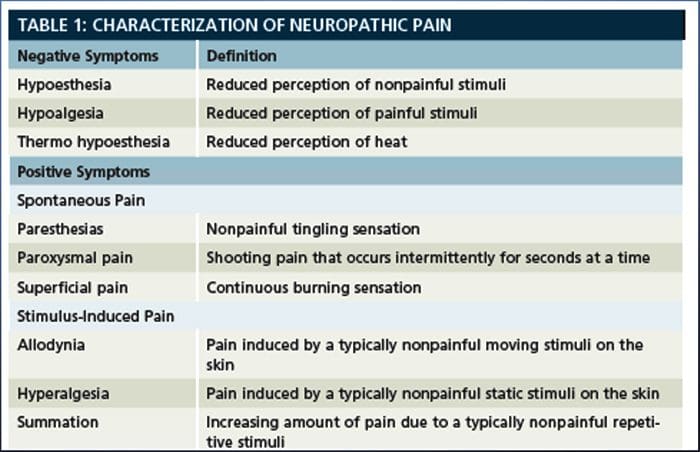
 PATHOGENESIS AF NEUROPATISK PAIN
PATHOGENESIS AF NEUROPATISK PAIN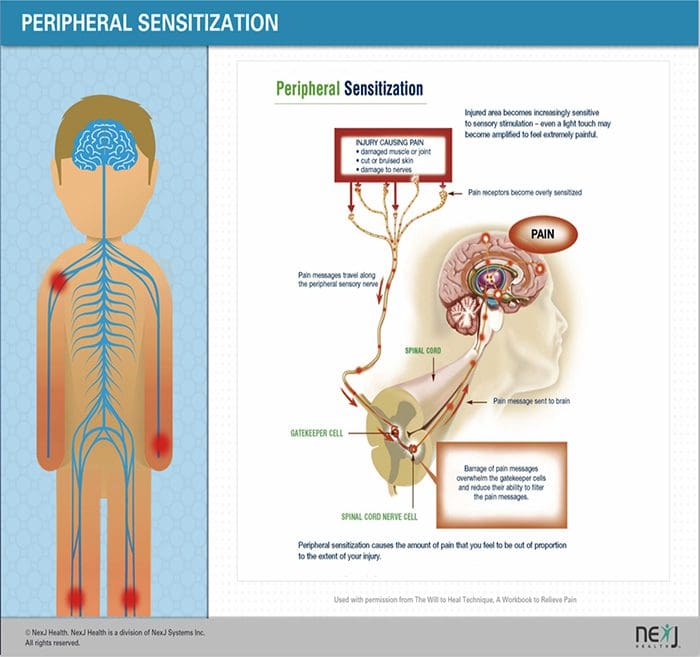
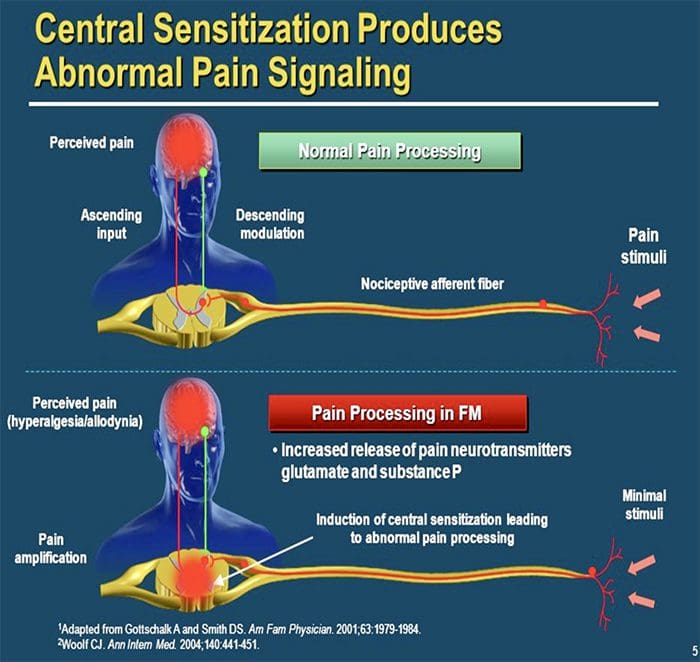
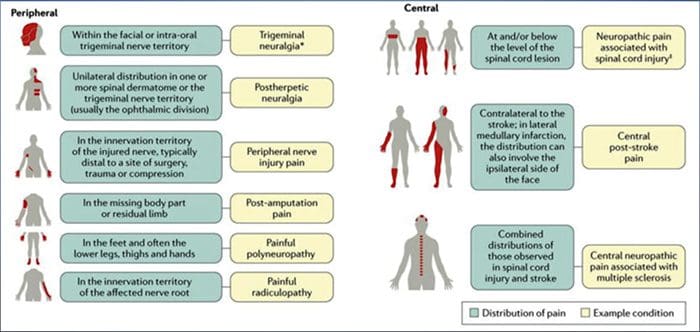 Almindelige årsager
Almindelige årsager
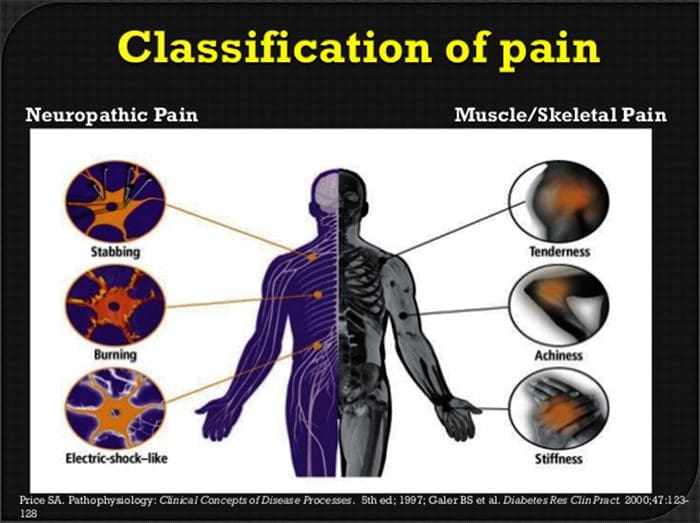
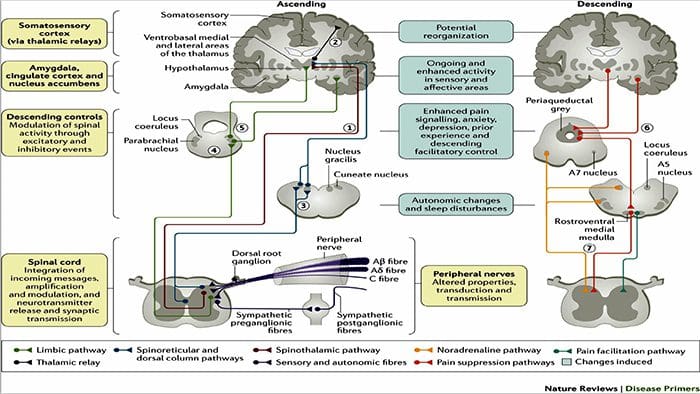
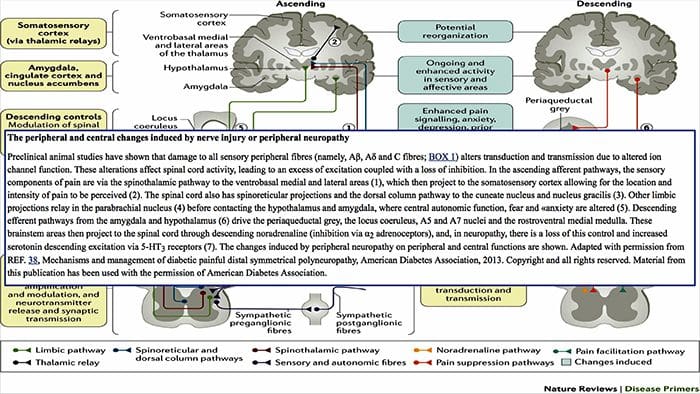

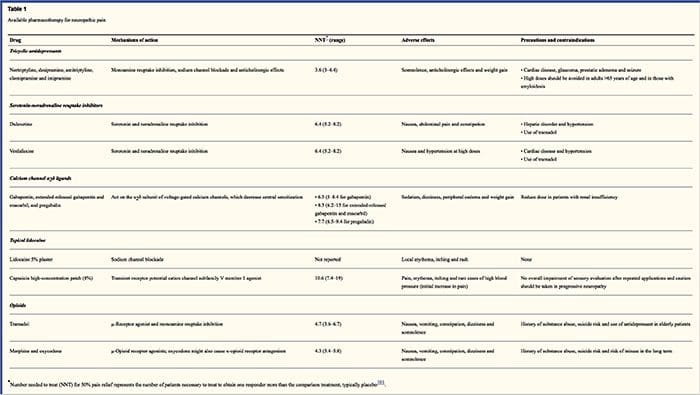




 PHANTOM LEMMESMÆRKE & AUSTERET VIRKELIGHED
PHANTOM LEMMESMÆRKE & AUSTERET VIRKELIGHED
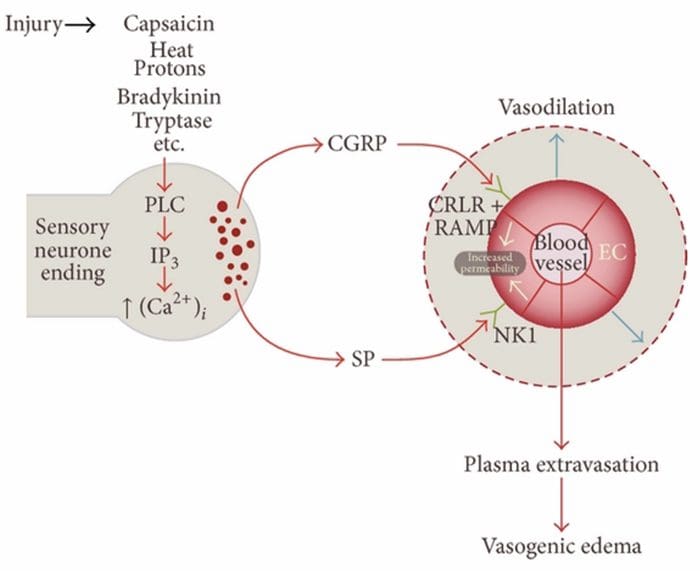 Almindelige årsager
Almindelige årsager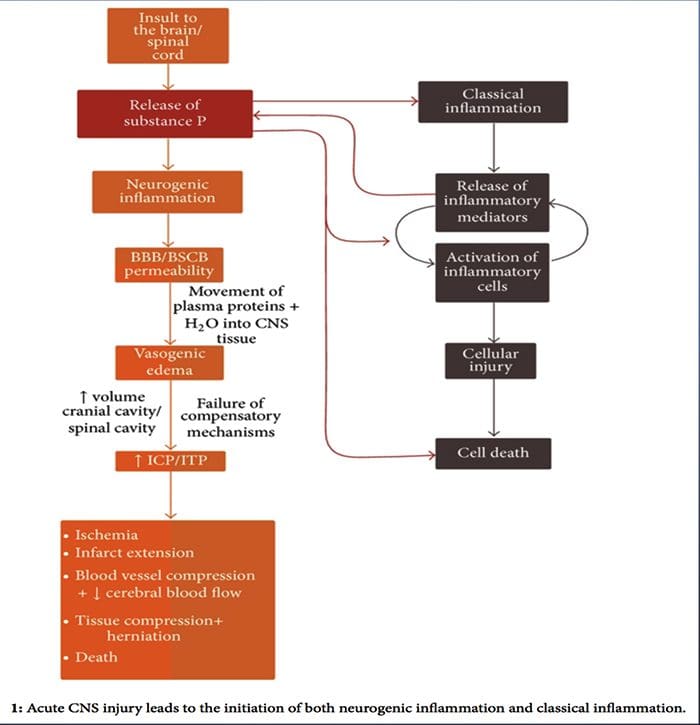


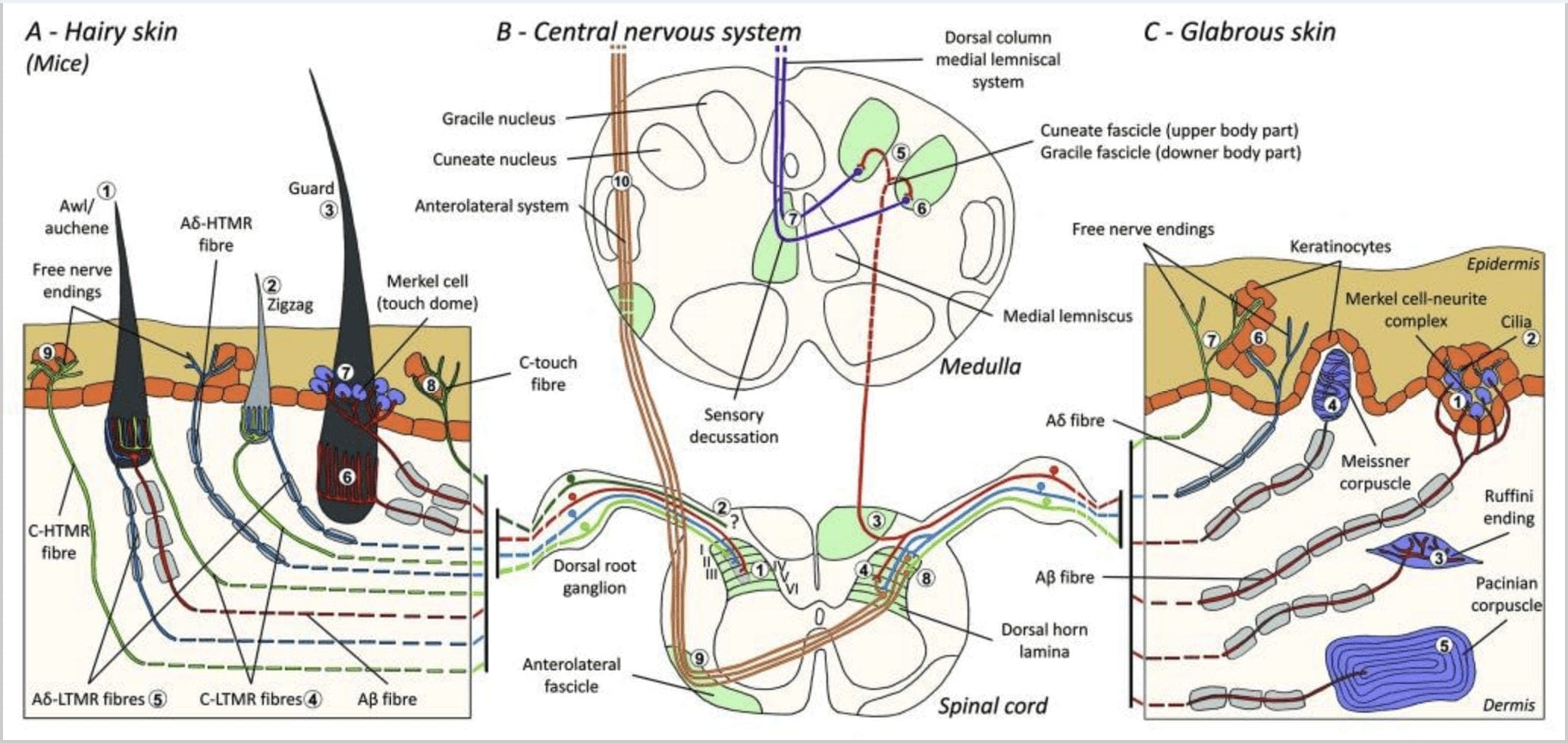
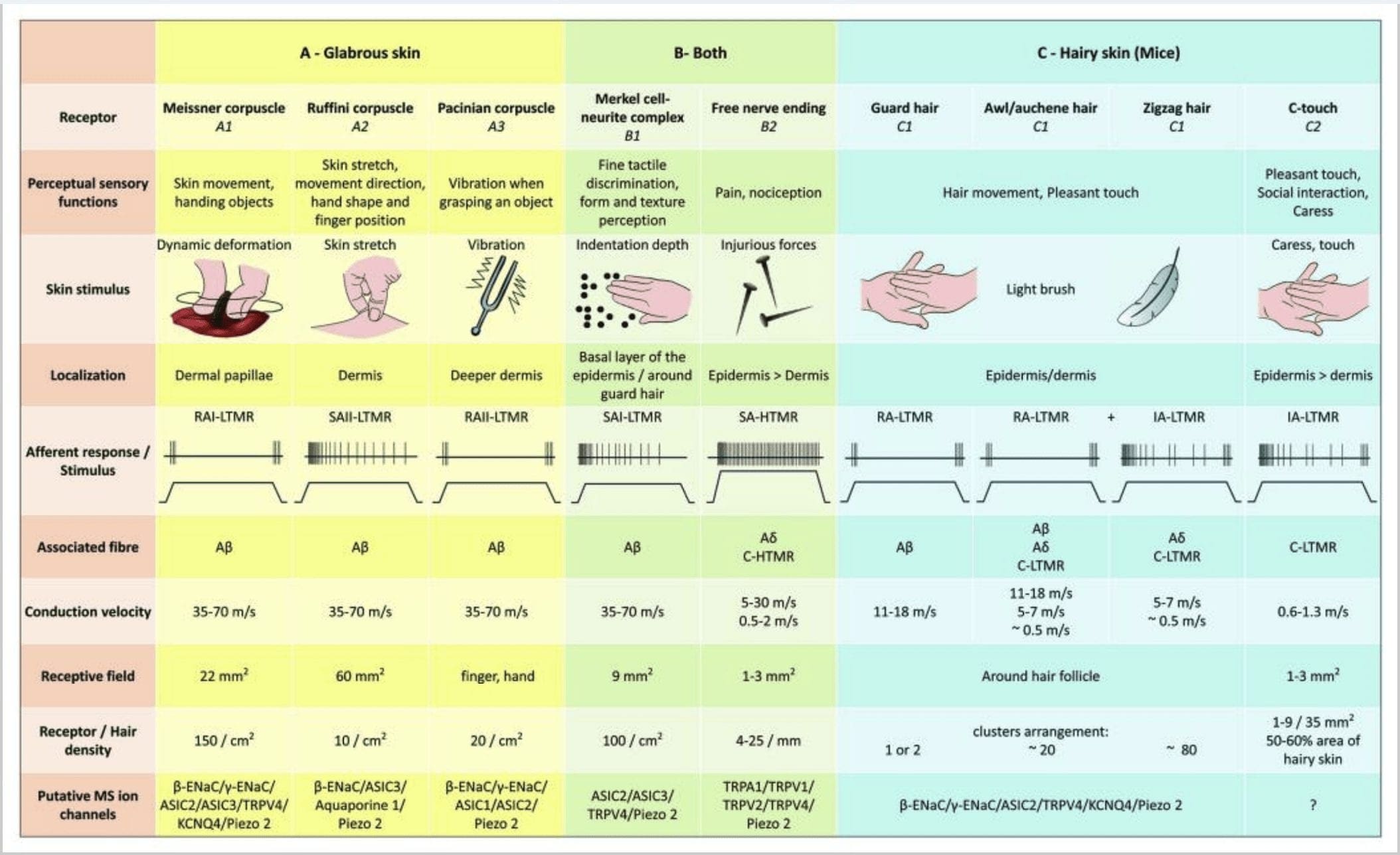

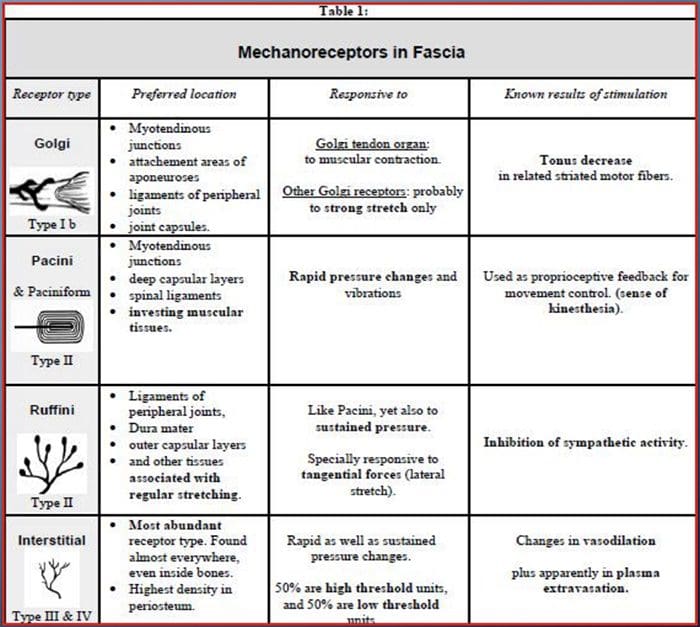 GATE KONTROL TEORI OM SMERTE
GATE KONTROL TEORI OM SMERTE
 DORSALSØLE MEDIAL LEMNISKAL VEJ
DORSALSØLE MEDIAL LEMNISKAL VEJ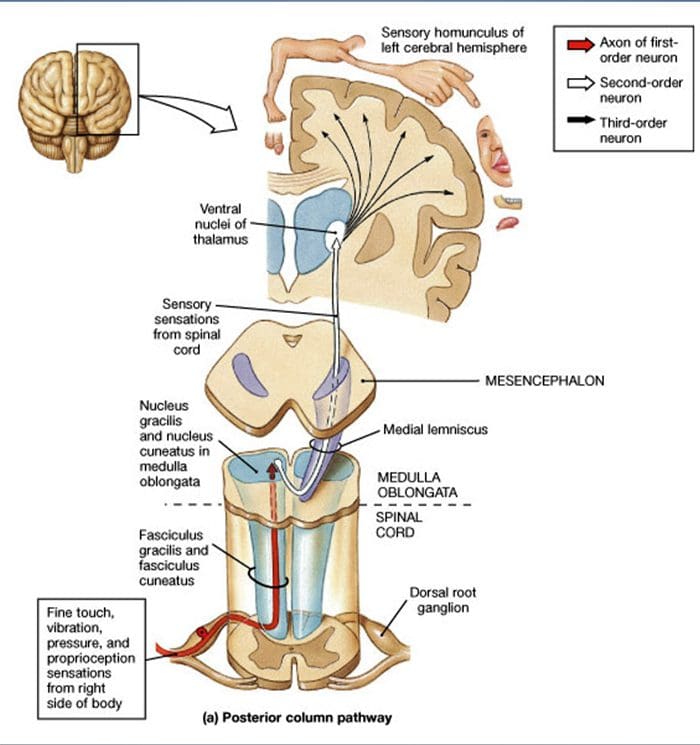 UDNYTTELSE AF PERIFERE MEKANORECEPTORER TIL AT ÆNDRE SMERTEPERCEPTION
UDNYTTELSE AF PERIFERE MEKANORECEPTORER TIL AT ÆNDRE SMERTEPERCEPTION
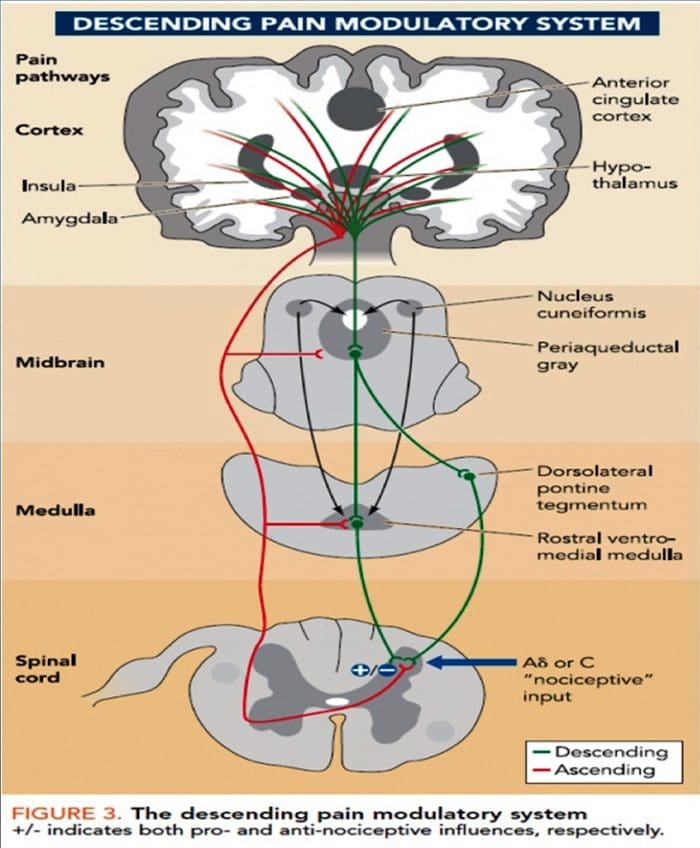
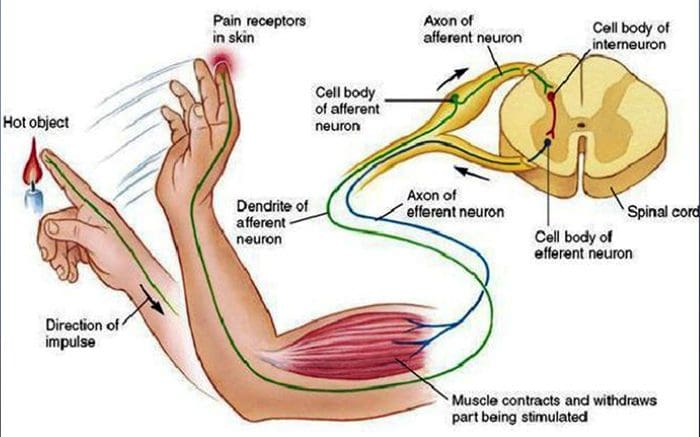
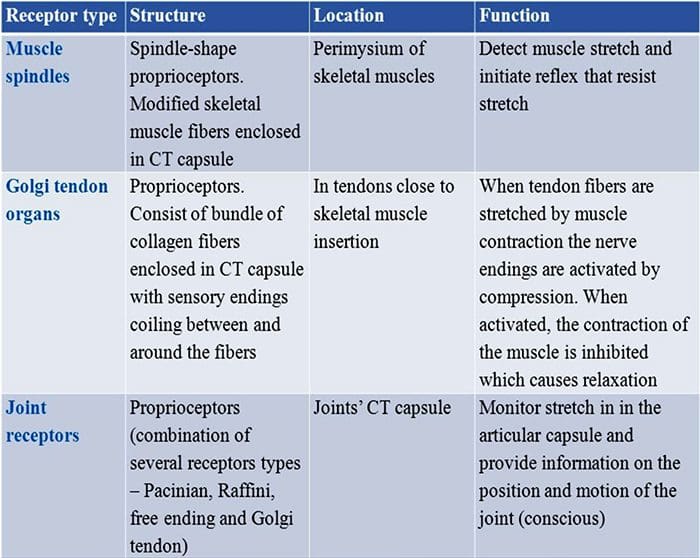
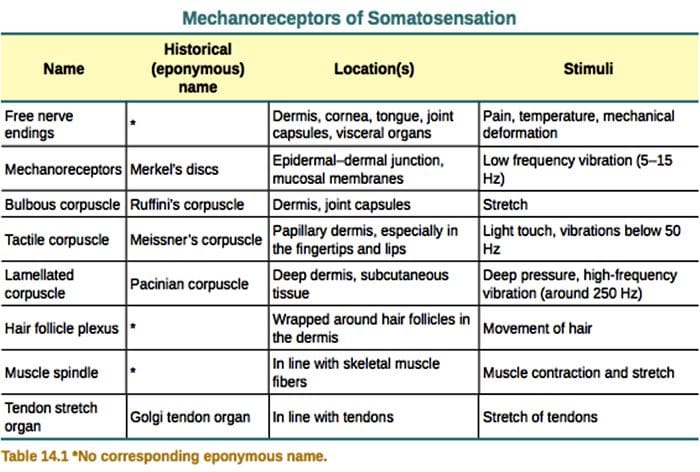 VIBRATION
VIBRATION
 LET BØR
LET BØR



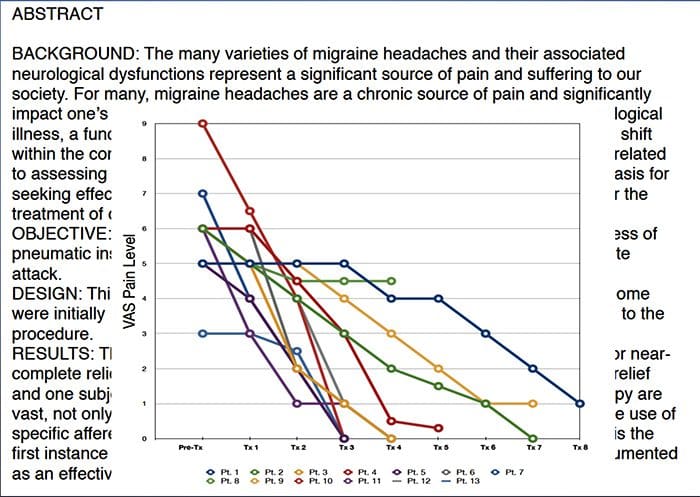


 CASESTUDIE
CASESTUDIE FYSISK UNDERSØGELSE HØJDEPUNKTER
FYSISK UNDERSØGELSE HØJDEPUNKTER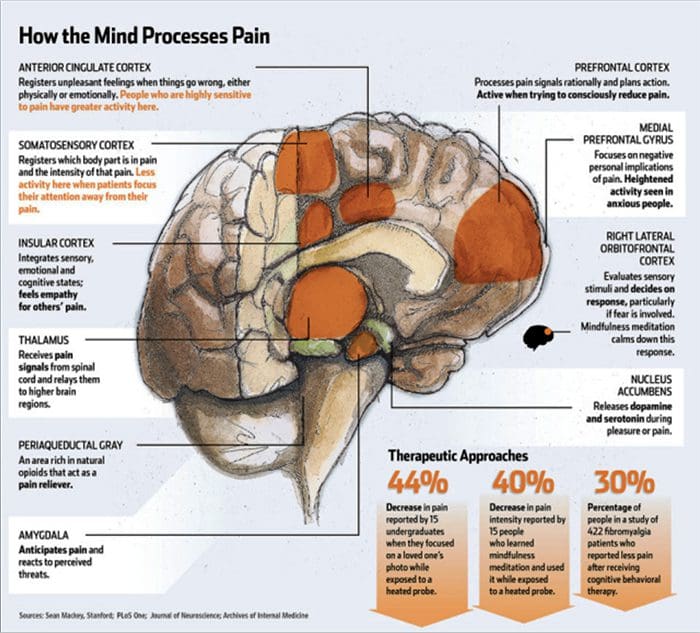 FYSISK UNDERSØGELSE HØJDEPUNKTER
FYSISK UNDERSØGELSE HØJDEPUNKTER
 EFTER TO DAGE
EFTER TO DAGE