Back Clinic Anti Aging Kiropraktik og Funktionel Medicin Team. Vores krop er i en konstant og uendelig kamp for overlevelse. Celler fødes, celler ødelægges. Forskere vurderer, at hver celle skal modstå over 10,000 individuelle angreb fra reaktive oxygenarter (ROS) eller frie radikaler. Uden fejl har kroppen et utroligt system af selvhelbredelse, der modstår angrebet og genopbygger det, der er blevet beskadiget eller ødelagt. Dette er skønheden i vores design.
At forstå aldringsbiologien og omsætte videnskabelig indsigt til interventioner, der forbedrer det sene liv gennem behandlinger. Det er nyttigt at have et klart, konsensussyn på, hvad der præcist udgør anti-aging behandling.
Siden før dagene med Ponce de Leons søgen efter lang levetid, har mennesket altid været lokket af chancen for evig ungdom. Kiropraktisk pleje med dens sundhedsbevægelse er en kraftfuld metode til at stabilisere og forbedre denne selvhelbredende evne. Dr. Alex Jimenez diskuterer koncepter omkring anti-aging pandora.
For personer, der ønsker at forbedre eller bevare hudens sundhed, kan inkorporering af akupunktur hjælpe med at forbedre huden og bekæmpe ældningsprocessen?
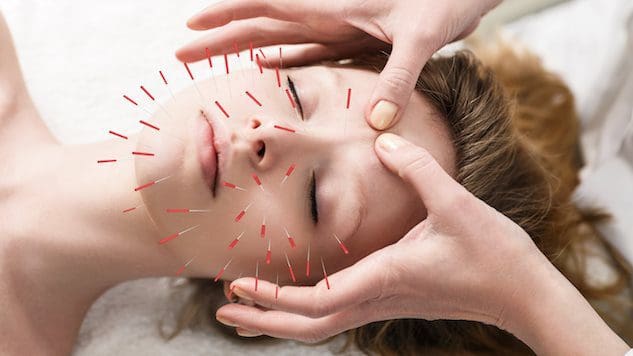
Kosmetisk akupunktur
Kosmetisk akupunktur følger den traditionelle akupunkturpraksis med nåleindføring. Målet er at vende ældningstegn og forbedre hudens sundhed. Det er nogle gange omtalt som akupunktur ansigtsforyngelse, som er blevet brugt som et alternativ til kirurgiske ansigtsløftninger og andre konventionelle procedurer. Foreløbige undersøgelser har undersøgt, hvordan det kan hjælpe med at fjerne alderspletter, løfte hængende øjenlåg og reducere rynker. (Younghee Yun et al., 2013)
Hvordan akupunktur virker
I traditionel kinesisk medicin eller TCM har akupunktur længe været brugt til at forbedre strømmen af energi – qi eller chi – i hele kroppen. Denne energi menes at cirkulere gennem energibaner kendt som meridianer. Når der opstår helbredsproblemer, er der ifølge TCM forhindringer eller blokeringer i kredsløbet.
Akupunktører kan genoprette optimal cirkulation/flow og forbedre sundheden ved at indsætte nåle i specifikke akupunkter. (National Institutes of Health, 2007)
Kosmetisk akupunktur
Kosmetisk akupunktur siges at forbedre hudens sundhed og fungere som en anti-aging behandling ved at stimulere produktionen af kollagen. Dette protein er en vigtig bestanddel af huden. Hudens indre lag mister kollagen og fasthed efterhånden som kroppen ældes. Der er dog behov for yderligere forskning for at understøtte påstanden om, at akupunktur kan fremme kollagenproduktionen. Nogle foreslår, at kosmetisk akupunktur hjælper med at forynge huden ved at forbedre kroppens samlede energi. En undersøgelse viste, at individer så forbedringer efter fem sessioner med kosmetisk ansigtsakupunktur. (Younghee Yun et al., 2013) Det anbefales dog, at der udføres ti behandlinger en eller to gange om ugen for at opnå optimale resultater. Derefter udføres vedligeholdelsesbehandlinger hver fjerde til ottende uge. I modsætning til Botox eller dermal fillers, er kosmetisk akupunktur ikke en hurtig løsning. Fokus er at skabe langsigtede forandringer i hud og krop, hvilket betyder forbedrede:
Når nålene sættes ind i huden, skaber de sår kendt som positive mikrotraumer. Kroppens naturlige helbredende og reparerende evner aktiveres, når den fornemmer disse sår. Disse punkteringer stimulerer lymfe- og kredsløbssystemerne, som leverer næringsstoffer og ilt til hudcellerne og nærer dem indefra og ud.
Dette hjælper med at udjævne teint og fremmer hudens udstråling.
De positive mikrotraumer stimulerer også produktionen af kollagen.
Dette hjælper med at forbedre elasticiteten, minimere linjer og rynker.
Alternativer
Adskillige naturlige midler kan hjælpe med at forbedre hudens sundhed og tilbyde anti-aging fordele. Ceramider er et fedtmolekyle, der findes naturligt i det øverste lag af huden og en ingrediens, der bruges i hudplejeprodukter. Disse kan beskytte mod ældningsrelateret tørhed i huden. (L Di Marzio 2008) Foreløbig forskning tyder på, at påføring af hvid te på huden kan bekæmpe nedbrydningen af kollagen og elastin – et protein, der understøtter hudens elasticitet og forhindrer nedbrydning). Der er også beviser for, at naturlige stoffer såsom arganolie, borageolie og havtorn kan tilbyde fugtgivende fordele, der kan forbedre huden.(Tamsyn SA Thring et al., 2009)
Selvom der er behov for yderligere beviser for kosmetisk akupunktur, kan integration af akupunktur hjælpe med at håndtere stress og forbedre det generelle helbred. Personer, der overvejer kosmetisk akupunktur, bør konsultere deres primære sundhedsplejerske for at se, om det er det rigtige for dem.
Forbedring af sundhed sammen: Omfavnelse af tværfaglig evaluering og behandling
Referencer
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS, & Choi, I. (2013). Effekt af ansigts kosmetisk akupunktur på ansigtets elasticitet: et åbent, enkeltarms pilotstudie. Evidensbaseret komplementær og alternativ medicin: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Nationalt Center for Komplementær og Alternativ Medicin. (2007). Akupunktur: en introduktion. Nationalt Center for Komplementær og Alternativ Medicin hjemmeside. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021). Reliability and Validity of Facial Check Sheet (FCS): Tjekliste for selvtilfredshed med kosmetisk akupunktur. Medicin (Basel, Schweiz), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Forøgelse af hudceramidniveauer hos ældre forsøgspersoner efter en kortvarig topisk applikation af bakteriel sphingomyelinase fra Streptococcus thermophilus. Internationalt tidsskrift for immunopatologi og farmakologi, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009). Anti-collagenase, anti-elastase og antioxidant aktiviteter af ekstrakter fra 21 planter. BMC komplementær og alternativ medicin, 9, 27. doi.org/10.1186/1472-6882-9-27
At holde en persons rygsøjle i topform er lig med mindre smerte og mere mobilitet, fleksibilitet og frihed. Kroppen slides ned og er en naturlig effekt af aldring, der sker for hver enkelt af os. Rygmarvsproblemer relateret til aldring kan blive alvorlige, hvis de ikke løses og vedtages med øvelser, udstrækning og kiropraktisk vedligeholdelse.
Aldring og ryggen
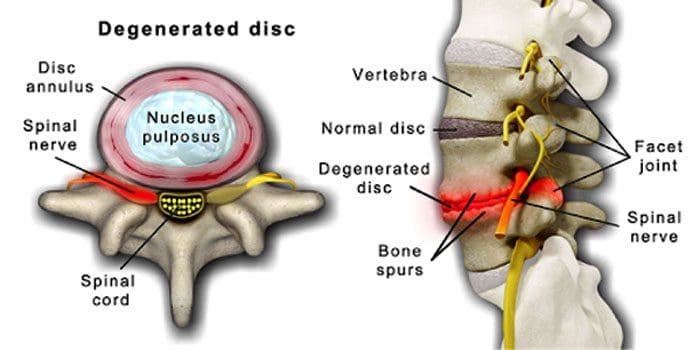
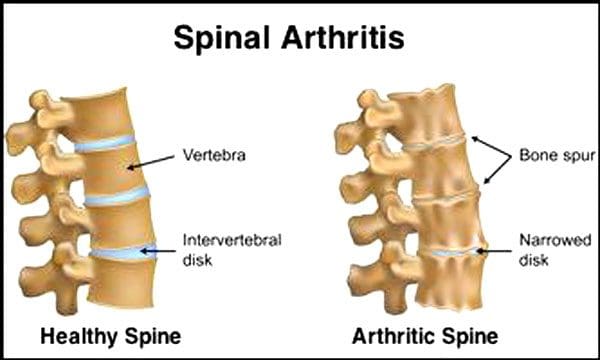
Det er normalt, at rygsøjlens diske og led forringes med alderen. Spinal stenose eller forsnævring af rygmarvskanalen kan også være en del af aldringsprocessen. To forhold forårsaget af aldring er degenerativ disk sygdom , artritis der også kan omfatte afstivning af rygsøjlens ledbånd og osteoporose.
Degenerativ diskussygdom opleves af 40% af personer i alderen 40 år
Stiger til 80 % for personer på 80 år og ældre.
Det centrerer sig omkring skiver, der gradvist skifter fra at være mest vand til mest fedt.
Når det er fedt, bliver skiverne indsnævret og mister elasticiteten.
Det siger Centers for Disease Control and Prevention 23% af amerikanske voksne har gigt. Dette er en tilstand, der hovedsageligt påvirker facetleddene. Leddene bliver hævede, hvilket reducerer bevægelsesområdet og kan påvirke spinalnerverne, hvilket forårsager smerte, svaghed og iskias. Med tiden stivner ledbåndene omkring og i rygsøjlen, hvilket reducerer bevægelsesområdet, hvilket forårsager stenose. Knogletab, eller osteoporose, er forårsaget af ændringer i hormoner og andre faktorer som ernæring. Aldring er en naturlig proces, men individer kan hjælpe deres rygsøjler med at forblive i topform, uanset hvor gamle de er.
Øvelse af sund kropsholdning
Lige fra hånden ordentlig sund kropsmekanik er et must. At forblive opmærksom og opmærksom på kropsholdningen bevarer justering og holder kroppen i balance. Sund kropsholdning vil hjælpe med at reducere virkningerne af:
Spinal stenose
Degenerativ disk sygdom
herniation
Risiko for rygmarvsbrud
At øve korrekt kropsholdning omfatter:
Reducer nedslidning
Sørg for, at arbejdsstationen er i topform og ergonomisk forsvarlig
Sørg for at bøje knæene, når du løfter, og hold rygsøjlen så lodret som muligt.
Yoga
Yoga kan være yderst gavnlig for en sundere, mere ungdommelig rygsøjle. Yoga opfylder tre områder for at holde rygsøjlen i topform. Dette inkluderer:
Regelmæssig motion
Bevarer fleksibiliteten
Opnår ideel kropsvægt
Yoga er en aldersforagtende aktivitet for rygsøjlen. Fordi det:
Bevarer styrken
Fleksibilitet
Posture
Balance
Kan være nyttigt for en række forskellige rygsygdomme, især gigtsmerter
Fald kan forårsage alvorlige skader. Yoga kan også hjælpe med at arbejde på balancen.
Se en kiropraktor
Forebyggende medicin er nøglen til at holde kroppen sund, ungdommelig og så stærk som muligt. En kiropraktisk undersøgelse kan afgøre, om der er rygmarvsproblemer og en diagnose for at udvikle en optimal behandlingsplan. Hvis kropsfunktionen er begrænset på grund af smerter i ryg og/eller ben, kontakt Injury Medical Kiropraktik og Funktionel Medicinsk Klinik og få rygsøjlen tilbage i topform.
Kropssammensætning
Motion/Stabilitet Ball Curls
Denne øvelse arbejder muskelgrupper, der er specifikke for spinal styrke og inkluderer:
Hamstrings
glutes
Dybe mavemuskler
Hoftebortførere og rotatorer
Øvelser som denne er en af de mest effektive måder at opbygge funktionel styrke og udholdenhed i baglår, hofter og forebygge skader. Sådan laver du denne træning:
Læg dig på ryggen med bøjede knæ
Løft benene op, så bunden af fødderne hviler oven på en træningsbold
Rul dine ben ud, indtil de er lige
Hold stillingen i et sekund eller to
Vend tilbage til toppen af bevægelsen, mens du klemmer hamstrings
Arbejde med disse muskler vil hjælpe med at gøre hugsiddende, longerende eller bøjede bevægelser lettere på rygsøjlen.
Ansvarsfraskrivelse for Dr. Alex Jimenez's blogindlæg
Omfanget af vores information er begrænset til kiropraktik, bevægeapparat, fysisk medicin, velvære og følsomme sundhedsmæssige problemer og / eller artikler om funktionel medicin, emner og diskussioner. Vi bruger funktionelle sundheds- og wellnessprotokoller til at behandle og understøtte pleje af skader eller lidelser i bevægeapparatet. Vores indlæg, emner, emner og indsigt dækker kliniske forhold, emner og emner, der relaterer og understøtter direkte eller indirekte vores kliniske anvendelsesområde. *
Vores kontor har gjort et rimeligt forsøg på at give støttende citater og har identificeret den eller de relevante forskningsundersøgelser, der understøtter vores stillinger. Vi stiller også kopier af støtteundersøgelser til rådighed for bestyrelsen og / eller offentligheden efter anmodning. Vi forstår, at vi dækker forhold, der kræver en yderligere forklaring på, hvordan det kan hjælpe i en bestemt plejeplan eller behandlingsprotokol; Derfor er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900 for yderligere at diskutere emnet ovenfor. Udbyderen (e) Licenseret i Texas og New Mexico *
De fødevarer, vi spiser, kan have potentiale til at være gavnlige eller skadelige for vores helbred. Dårlig ernæring kan forårsage en række sundhedsproblemer, herunder fedme, hjerte-kar-sygdomme og type 2-diabetes. I mellemtiden kan korrekt ernæring få dig til at føle dig energisk, reducere din risiko for helbredsproblemer samt hjælpe med at opretholde og regulere en sund vægt. Hvis du vil fremme lang levetid, skal du brænde din krop med god mad. I den følgende artikel vil vi liste flere gode fødevarer, der i sidste ende kan hjælpe med at fremme lang levetid ved også at hjælpe med at forbedre den generelle sundhed og velvære.
Krydderige Grøntsager
Korsblomstrede grøntsager har den unikke evne til at ændre vores hormoner, udløse kroppens naturlige afgiftningssystem og endda reducere væksten af kræftceller. Disse skal tygges grundigt eller spises strimlet, hakket, juicet eller blandet for at frigive deres gavnlige egenskaber. Sulforaphane, der findes i korsblomstrede grøntsager, har også vist sig at hjælpe med at beskytte blodkarvæggen mod betændelse, der kan forårsage hjertesygdomme. Korsblomstrede grøntsager, såsom grønkål, kål, rosenkål, blomkål og broccoli er flere af de mest næringsrige fødevarer i verden.
Grøn salat
Rå bladgrønt har mindre end 100 kalorier pr. pund, hvilket gør dem til den perfekte mad til vægttab. At spise mere grønt salat har også været forbundet med den reducerede risiko for hjerteanfald, slagtilfælde, diabetes og flere typer kræft. Rå bladgrønt er også rige på det essentielle B-vitamin folat, plus lutein og zeaxanthin, carotenoider, der kan hjælpe med at beskytte øjnene. Fedtopløselige fytokemikalier, såsom carotenoider, der findes i salatgrønt som salat, spinat, grønkål, collard greens og sennepsgrønt har også antioxidante og antiinflammatoriske virkninger i kroppen.
Nødder
Nødder er en lavglykæmisk mad og en fantastisk kilde til sunde fedtstoffer, planteprotein, fibre, antioxidanter, phytosteroler og mineraler, hvilket også hjælper med at reducere den glykæmiske belastning af et helt måltid, hvilket gør dem til en væsentlig del af en anti-diabetes kost. Uanset deres kalorietæthed kan det at spise nødder hjælpe med at fremme vægttab. Nødder kan også reducere kolesterol og hjælpe med at reducere risikoen for hjertesygdomme.
Frø
Frø, ligesom nødder, giver også sunde fedtstoffer, antioxidanter og mineraler, men disse har mere protein og er rige på spormineraler. Chia-, hør- og hampefrø er rige på omega-3 fedtstoffer. Chia-, hør- og sesamfrø er også rige lignaner eller fytoøstrogener, der bekæmper brystkræft. Desuden er sesamfrø rige på calcium og vitamin E, og græskarfrø er rige på zink.
Bær
Bær er antioxidantrige frugter, der kan hjælpe med at fremme hjertesundheden. Forskningsundersøgelser, hvor deltagerne spiste jordbær eller blåbær dagligt i flere uger, rapporterede forbedringer i blodtryk, total- og LDL-kolesterol og endda tegn på oxidativt stress. Bær har også anti-kræft egenskaber og har vist sig at hjælpe med at forhindre kognitiv tilbagegang forbundet med aldring.
Granatæble
Den mest kendte fytokemikalie i granatæbler, punicalagin, er ansvarlig for mere end halvdelen af frugtens antioxidantaktivitet. Granatæblefytokemikalier har anti-kræft, hjertebeskyttende og hjernesunde fordele. I en forskningsundersøgelse klarede ældre voksne, der drak granatæblejuice dagligt i 28 dage, bedre på en hukommelsestest sammenlignet med dem, der drak en placebo-drik.
Bønner
At spise bønner og andre bælgfrugter kan hjælpe med at balancere blodsukkeret, reducere din appetit og beskytte mod tyktarmskræft. Bønner er en anti-diabetes fødevare, der kan hjælpe med at fremme vægttab, fordi de fordøjes langsomt, hvilket bremser stigningen i blodsukkeret efter et måltid og hjælper med at forhindre madtrang ved at fremme mæthed. At spise bønner og andre bælgfrugter to gange om ugen har vist sig at mindske risikoen for tyktarmskræft. At spise bønner og andre bælgfrugter, såsom røde bønner, sorte bønner, kikærter, linser og flækærter, giver også betydelig beskyttelse mod andre kræftformer.
Svampe
Regelmæssig spisning af svampe er forbundet med en reduceret risiko for brystkræft. Hvide og Portobello-svampe er især gavnlige mod brystkræft, fordi de har aromatasehæmmere eller forbindelser, der hæmmer produktionen af østrogen. Svampe har vist sig at have anti-inflammatoriske virkninger samt give øget immuncelleaktivitet, forebyggelse af DNA-skader, bremset kræftcellevækst og angiogenesehæmning. Svampe bør altid koges, da rå svampe har et potentielt kræftfremkaldende kemikalie kendt som agaritin, der reduceres betydeligt ved tilberedning.
Løg og hvidløg
Løg og hvidløg giver kardiovaskulære og immunsystem fordele samt giver anti-diabetiske og anti-cancer effekter. Disse har også været forbundet med en lavere risiko for mave- og prostatacancer. Løg og hvidløg er kendt for deres organiske svovlforbindelser, som hjælper med at forhindre udviklingen af kræft ved at afgifte kræftfremkaldende stoffer, reducere kræftcellevækst og blokere angiogenese. Løg og hvidløg har også høje koncentrationer af sundhedsfremmende flavonoid-antioxidanter, som har antiinflammatoriske virkninger, der kan være med til at forebygge kræft.
tomater
Tomater er rige på en række forskellige næringsstoffer, såsom lycopen, vitamin C og E, beta-caroten og flavonol antioxidanter. Lycopen kan hjælpe med at beskytte mod prostatacancer, UV-hudskader og? kardiovaskulær sygdom. Lycopen absorberes bedre, når tomater koges. En kop tomatsauce har omkring 10 gange mængden af lycopen som en kop rå, hakkede tomater. Husk også på, at carotenoider, ligesom lycopen, absorberes bedst, når de ledsages af sunde fedtstoffer, så nyd dine tomater i en salat med nødder eller en nøddebaseret dressing for ekstra ernæringsmæssige fordele.
De fødevarer, vi spiser, kan have potentiale til at være gavnlige eller skadelige for vores helbred. Dårlig ernæring kan forårsage en række sundhedsproblemer, herunder fedme, hjerte-kar-sygdomme og type 2-diabetes. I mellemtiden kan korrekt ernæring få dig til at føle dig energisk, reducere din risiko for helbredsproblemer samt hjælpe med at opretholde og regulere en sund vægt. Hvis du vil fremme lang levetid, skal du brænde din krop med god mad. God mad kan også hjælpe med at reducere betændelse forbundet med en række sundhedsproblemer, herunder ledsmerter og gigt. Sundhedspersonale, såsom kiropraktorer, kan tilbyde kost- og livsstilsrådgivning for at hjælpe med at fremme sundhed og velvære. I den følgende artikel vil vi liste flere gode fødevarer, der i sidste ende kan være med til at fremme lang levetid. – Dr. Alex Jimenez DC, CCST Insight
Zesty Beet Juice
Portioner: 1 Tilberedningstid: 5-10 minutter
1 grapefrugt, skrællet og skåret
1 æble, vasket og skåret
1 hel roer og efterlader, hvis du har dem, vasket og skåret
1-tommers knop ingefær, skyllet, skrællet og hakket
Saft alle ingredienser i en saftpresser i høj kvalitet. Bedst tjent med det samme.
Bare en gulerod giver dig alt dit daglige vitamin A-indtag
Ja, at spise kun en kogt 80 g gulerod giver dig nok beta-caroten til din krop til at producere 2 mikrogram (mcg) A-vitamin (nødvendigt til fornyelse af hudceller). Det er mere end det anbefalede daglige indtag af A-vitamin i USA, som er omkring 1,480mcg. Det er bedst at spise kogte gulerødder, da dette blødgør cellevæggene, så mere beta-caroten absorberes. Tilføjelse af sundere fødevarer i din kost er en fantastisk måde at forbedre dit generelle helbred på.
Omfanget af vores information er begrænset til kiropraktik, bevægeapparat, fysisk medicin, wellness og følsomme sundhedsmæssige problemer og / eller funktionelle medicinske artikler, emner og diskussioner. Vi bruger funktionelle sundheds- og wellnessprotokoller til at behandle og understøtte pleje af skader eller lidelser i bevægeapparatet. Vores indlæg, emner, emner og indsigt dækker kliniske spørgsmål, emner og emner, der direkte eller indirekte relaterer til vores kliniske praksis. * Vores kontor har gjort et rimeligt forsøg på at give støttende citater og har identificeret den relevante forskningsundersøgelse eller undersøgelser, der understøtter vores stillinger. Vi stiller også kopier af understøttende forskningsstudier til rådighed for bestyrelsen og / eller offentligheden efter anmodning. Vi forstår, at vi dækker forhold, der kræver en yderligere forklaring på, hvordan det kan hjælpe i en bestemt plejeplan eller behandlingsprotokol; derfor, for yderligere at diskutere emnet ovenfor, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900. Udbyderen (e) Licenseret i Texas * og New Mexico *
Kurateret af Dr. Alex Jimenez DC, CCST
Referencer:
Joel Fuhrman, MD. �10 bedste fødevarer, du kan spise for at leve længere og forblive sund.� Verywell Health, 6. juni 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. Kaffe er en frugt og andre utroligt sande madfakta. MSN livsstil4. juni 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Hvis du oplever nogen af disse situationer, kan dine kollagenpeptider være lave.
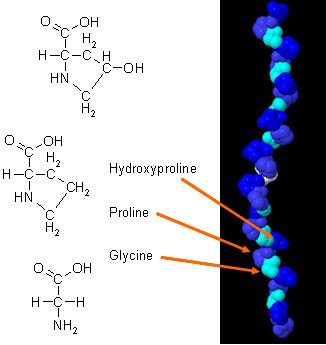
Der har været nye undersøgelser om hvordan kollagen kan forbedre kropssammensætningen, når det kombineres med daglige øvelser. Kollagen i kroppen har en unik aminosyresammensætning, der spiller en væsentlig rolle i kroppens anatomi. Kollagenprotein er en koncentreret kilde til glycin, prolin og hydroxyprolin, og når det sammenlignes med alle de andre diætproteiner, gør det kollagen til et potentielt praktisk valg som et strukturelt protein.
In en 2015 undersøgelse, har forskere demonstreret, hvordan effektive kollagentilskud kan forbedre kropssammensætningen hos aktive mænd. Resultaterne viser, hvordan hver mandlig individ deltager i vægttræning mindst tre gange om ugen og skal supplere med mindst 15 gram kollagenpeptider for at opnå maksimal sundhed. De vurderinger, som testen giver, er styrketest, bioimpedansanalyse (BIA) og muskelbiopsier. Disse tests sikrer, at de mandlige individer klarer sig godt efter at have taget kollagentilskud, og resultaterne viser, hvordan deres kropsmasse havde en stigning i fedtfri kropsmasse. En anden undersøgelse viste, hvordan tilskud af kollagenprotein, når det kombineres med styrketræning, kan øge muskelmasse og muskelstyrke hos ældre såvel som personer med sarkopeni.
Fordelagtige egenskaber med kollagen
Der er mange gavnlige egenskaber at kollagentilskud kan give til kroppen, når det indtages. Der er hydrolyseret kollagen og gelatine og kan hjælpe med at forbedre en persons hudstruktur. Selvom der ikke er mange undersøgelser om kollagentilskud, er der gode løfter for områderne på kroppen. De er:
Muskelmasse: Kollagentilskud kan, når det kombineres med styrketræning, øge muskelmasse og styrke i kroppen.
Artritis: Kollagentilskud kan hjælpe folk med slidgigt. Undersøgelser viser at når folk slidgigt tager kollagentilskud, opdagede de et massivt fald i den smerte, de oplevede.
Hudens elasticitet: I en 2014 undersøgelse, det erklærede, at kvinder, der tog kollagen kosttilskud og har vist forbedringer i hudens elasticitet. Kollagen kan også bruges i topiske behandlinger for at hjælpe med at forbedre udseendet af en persons hud ved at minimere fine linjer og rynker.
Ikke kun kollagentilskud giver gavnlige egenskaber til de specifikke områder på kroppen, men der er de fire hovedtyper af kollagen, og hvad er deres roller i den menneskelige krop såvel som deres funktioner:
Type 1: Type 1 kollagen tog højde for 90 % af kroppens kollagen og består af tætpakkede fibre, der giver strukturer til huden, knoglerne, bindevævet og tænderne i kroppen.
Type 2: Type 2 kollagen er opbygget af løst pakkede fibre, der findes i den elastiske brusk, som hjælper med at polstre leddene i kroppen.
Type 3: Type 3 kollagen hjælper med at understøtte strukturen af musklerne, organerne og arterierne, der sørger for, at kroppen fungerer korrekt.
Type 4: Type 4 kollagen findes i lagene af alles hud og hjælper med filtreringen i kroppen.
Da disse fire typer af kollagen er i kroppen, er det vigtigt at vide, at kollagen naturligt kan falde over tid med alderen, da kroppen vil producere en mindre, lavere kvalitet af kollagen. Et af de synlige tegn på fald i kollagen er, når huden på den menneskelige krop bliver mindre fast og smidig samt svækker brusk på grund af aldring.
Faktorer, der kan skade kollagen
Selvom kollagen kan falde naturligt med alderen, kan mange faktorer ødelægge kollagener, der er skadelige for huden. De skadelige faktorer kan omfatte:
Sukker og kulhydrater: Raffinerede sukkerarter og kulhydrater kan forstyrre med kollagens evne til at reparere sig selv på huden. Så ved at minimere sukker- og kulhydratforbruget i kroppen kan det reducere virkningerne af vaskulær, nyre- og kutanvævsdysfunktion.
Soleksponering: Selvom det at få nok sol kan hjælpe en person med at nyde dagen, men at blive udsat for solen i en længere periode kan forårsage beskadigede til huden og ødelægge kollagenpeptider. Virkningerne af overeksponering af solen kan få huden til at fotografere alder og producere oxidativ stress i kroppen.
Smoking: Når en person ryger, kan den reducere kollagenproduktionen i kroppen, hvilket gør, at kroppen får for tidlige rynker, og hvis kroppen er såret, vil helingsprocessen gå langsommere og kan føre til lidelser i kroppen.
autoimmune sygdomme: Nogle autoimmune sygdomme kan også skade kollagenproduktionen som lupus.
Konklusion
Kollagen er livsvigtigt for kroppen, da det hjælper huden med at være blid og fast. Naturligvis vil det falde, når en person bliver ældre, så at tage kollagentilskud kan sikre, at kroppen kan fungere korrekt. Når skadelige faktorer påvirker kroppen, kan de stoppe eller endda beskadige kollagenproduktionen og fremskynde processen med at danne for tidlige rynker, hvilket får en person til at se ældre ud, end den er. Nogle produkter kan hjælpe kroppens cellulære aktivitet ved at give mere fremragende stabilitet, biotilgængelighed og fordøjelseskomfort.
Omfanget af vores information er begrænset til kiropraktik, muskuloskeletale og nervesundhedsmæssige problemer eller artikler, emner og diskussioner om funktionel medicin. Vi bruger funktionelle sundhedsprotokoller til at behandle skader eller lidelser i bevægeapparatet. Vores kontor har gjort et rimeligt forsøg på at give støttende citater og har identificeret den relevante forskningsundersøgelse eller undersøgelser, der understøtter vores stillinger. Vi stiller også kopier af understøttende forskningsundersøgelser til rådighed for bestyrelsen og eller offentligheden efter anmodning. For yderligere at diskutere emnet ovenfor, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900.
Referencer:
Bosch, Ricardo, et al. �Mekanismer for fotoaldring og kutan fotokarcinogenese og fotobeskyttende strategier med fytokemikalier.� Antioxidanter (Basel, Schweiz), MDPI, 26. marts 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, F William. �Ernæring og aldrende hud: Sukker og glykering.� Klinikker i Dermatologi, US National Library of Medicine, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. � Kollagen – hvad er det, og hvad er det godt for?� Healthline, 9. september 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. �Ny undersøgelse demonstrerer fordelene ved kollagenpeptider til forbedring af kropssammensætning kombineret med motion.� Design til sundhed, 31. maj 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. �Rygning påvirker kollagensyntese og ekstracellulær matrixomsætning i menneskelig hud.� British Journal of Dermatology, US National Library of Medicine, april 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, et al. �Oralt tilskud af specifikke kollagenpeptider har gavnlige virkninger på menneskets hudfysiologi: en dobbeltblind, placebokontrolleret undersøgelse.� Hudfarmakologi og fysiologi, US National Library of Medicine, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, et al. �Effekten af det nye hydrolyserede kyllingebriskekstrakt med lav molekylvægt, BioCell Collagen, på forbedring af slidgigt-relaterede symptomer: et randomiseret, dobbeltblindt, placebokontrolleret forsøg.� Journal of Agricultural and Food Chemistry, US National Library of Medicine, 25 apr. 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise, et al. �Collagenpeptidtilskud i kombination med modstandstræning forbedrer kropssammensætning og øger muskelstyrken hos ældre sarkopeniske mænd: et randomiseret kontrolleret forsøg.� British Journal of Nutrition, Cambridge University Press, 28. oktober 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Ved at informere enkeltpersoner om, hvordan National University of Health Sciences leverer viden til fremtidige generationer, tilbyder universitetet en bred vifte af medicinske professioner til funktionel medicin.
Ligesom du er blevet diagnosticeret med cøliaki, irritabel tarmsyndrom, divertikulose / diverticulitis eller lækkende tarmsyndrom?
Overdreven bøjning, burping eller oppustethed?
Unormal distention efter visse probiotika eller naturlige kosttilskud?
Mistanke om ernæringsmæssig malabsorption?
Afbryder fordøjelsesproblemerne med afslapning?
Hvis du oplever nogen af disse situationer, oplever du muligvis tarmproblemer og bliver muligvis nødt til at prøve 4R-programmet.
Fødevarefølsomhed, reumatoid arthritis og angst har været forbundet med nedsat gastrointestinal permeabilitet. Disse forskellige tilstande kan ske fra mange faktorer, der kan påvirke fordøjelseskanalen. Hvis den ikke behandles, kan det muligvis være et resultat af dysfunktion i tarmpermeabilitetsbarrieren, der forårsager betændelse og alvorlige helbredsforhold, som tarmen kan udvikle. 4R-programmet bruges til at gendanne en sund tarm i kroppen og involverer fire trin. De er: fjerne, udskifte, genokuleret og reparere.
Intestinal permeabilitet
Tarmpermeabiliteten hjælper med at beskytte kroppen og sørger for, at skadelige bakterier ikke kommer ind i tarmen. Det beskytter kroppen mod potentielle miljøfaktorer det kan være skadeligt og kommer ind gennem fordøjelseskanalen. Det kan enten være toksin, patogene mikroorganismer og andre antigener, der kan skade fordøjelseskanalen og forårsage problemer. Tarmforingen består af et lag epitelceller, der er adskilt ved tætte forbindelser. I en sund tarm, regulerer det tætte knudepunkt tarmpermeabiliteten ved selektivt at tillade stoffer at komme ind og bevæge sig over tarmbarrieren og forhindre, at skadelige faktorer optages.
Visse miljøfaktorer kan beskadige det tætte kryds, og resultatet er, at det kan øge tarmpermeabiliteten, hvilket forårsager tarmens hyperpermeabilitet eller lækkende tarm i kroppen. Medvirkende faktorer kan øge tarmpermeabiliteten som en overdreven mængde mættet fedt og alkohol, mangler i næringsstoffer, kronisk stress og infektionssygdomme.
Med en forøget tarmpermeabilitet i tarmen kan det gøre det muligt for antigener at krydse tarmslimhinden og komme ind i blodbanen og forårsage et immunrespons og betændelse i kroppen. Der er visse gastrointestinale tilstande, der er forbundet med hyperpermeabilitet i tarmen, og hvis den ikke behandles, kan det udløse visse autoimmune tilstande, der kan forårsage skade på kroppen.
4Rs-program
4Rs er et program, som sundhedspersonale rådgiver deres patienter til at bruge, når de adresserer forstyrrende fordøjelsesproblemer og hjælper med at støtte tarmheling.
Fjernelse af problemet
Det første trin i 4Rs-programmet er at fjerne skadelige patogener og betændelsesudløsere, der er forbundet med øget tarmpermeabilitet. Udløsere som stress og kronisk alkoholforbrug kan skade en persons krop meget. Så at målrette mod disse skadelige faktorer fra kroppen er at behandle det med medicin, antibiotika, kosttilskud, og fjernelse af inflammatoriske fødevarer fra kosten anbefales, herunder:
- Alkohol
- Gluten
- Madtilsætningsstoffer
- Stivelse
- Visse fedtsyrer
- Visse fødevarer, som en person er følsom overfor
Udskiftning af næringsstoffer
Det andet trin i 4Rs-programmet er at erstatte de næringsstoffer, der forårsager tarmen problemer gennem betændelse. Visse næringsstoffer kan hjælpe med at reducere betændelse i tarmen, mens du sørger for, at fordøjelseskanalen understøttes. Der er nogle antiinflammatoriske fødevarer, der er næringsrige. Disse inkluderer:
- Fødevarer med meget fiber
- Omega-3
- Olivenolie
- Svampe
- Antiinflammatoriske urter
Der er visse kosttilskud, der kan bruges til at støtte fordøjelsesfunktionen ved at hjælpe og absorbere næringsstofferne til at fremme en sund tarm. Hvad fordøjelsesenzymerne gør, er at de hjælper med at hjælpe med at nedbryde fedt, proteiner og kulhydrater i tarmen. Dette vil hjælpe personer, der har en nedsat fordøjelseskanal, fødevareintolerancer eller har cøliaki, til gavn for dem. Tilskud som galdesyretilskud kan hjælpe med til absorption af næringsstoffer ved at flette lipider sammen. Undersøgelser har oplyst at galdesyrer er blevet brugt til behandling af leveren, galdeblæren og galdekanalen, mens de forhindrer dannelse af galdesten efter bariatrisk kirurgi.
Rejokuleret tarmen
Det tredje trin er af 4rs-programmet til genokulation af tarmen mikrob med gavnlige bakterier for at fremme en sund tarmsfunktion. Undersøgelser er vist at probiotiske kosttilskud er blevet brugt til at forbedre tarmen ved at gendanne gavnlige bakterier. Med disse kosttilskud giver de tarmen en forbedring ved at udskille antiinflammatoriske stoffer i kroppen, hjælpe med at støtte immunforsvaret, ændre kroppens mikrobielle sammensætning og reducere tarmpermeabiliteten i tarmsystemet.
Siden probiotika findes i gærede fødevarer og betragtes som en kortvarig, da de ikke er vedvarende i mave-tarmkanalen og er gavnlige. Overraskende har de stadig en indflydelse på menneskers sundhed på grund af indflydelse på tarmen ved at producere vitaminer og antimikrobielle forbindelser, hvilket giver mangfoldighed og tarmsfunktion.
Reparation af tarmen
Det sidste trin i 4Rs-programmet er at reparere tarmen. Dette trin involverer reparation af tarmens foring af tarmen med specifikke næringsstoffer og urter. Disse urter og kosttilskud kan hjælpe med at mindske tarmpermeabiliteten og betændelse i kroppen. Nogle af disse urter og kosttilskud inkluderer:
- Aloe vera
- Chios mastik gummi
- DGL (Deglycyrrhizinated lakrits)
- Marshmallow rod
- L-glutamin
- Omega-3
Polyphenoler
- D-vitamin
- Zink
Konklusion
Da mange faktorer kan have en negativ indvirkning på fordøjelsessystemet på en skadelig måde og kan være bidragyder til adskillige sundhedsmæssige tilstande. Hovedmålet med 4Rs-programmet er at minimere disse faktorer, der skader tarmen og reducerer betændelse og øget tarmpermeabilitet. Når patienten introduceres til de gavnlige faktorer, som 4R'erne leverer, kan det føre til en sund, helet tarm. Nogle produkter er her for at hjælpe med at understøtte mave-tarm-systemet ved at støtte tarmene, forbedre sukkermetabolismen og målrette mod aminosyrerne, der er beregnet til at støtte tarmene.
Omfanget af vores information er begrænset til kiropraktik, muskuloskeletale og nervesundhedsmæssige problemer eller artikler, emner og diskussioner om funktionel medicin. Vi bruger funktionelle sundhedsprotokoller til at behandle skader eller lidelser i bevægeapparatet. Vores kontor har gjort et rimeligt forsøg på at give støttende citater og har identificeret den relevante forskningsundersøgelse eller undersøgelser, der understøtter vores stillinger. Vi stiller også kopier af understøttende forskningsundersøgelser til rådighed for bestyrelsen og eller offentligheden efter anmodning. For yderligere at diskutere emnet ovenfor, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900.
Referencer:
De Santis, Stefania, et al. Næringsnøgler til tarmbarriermodulation . Grænser i immunologi, Frontiers Media SA, 7 dec. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca, et al. Supplering af fordøjelsesenzym ved mave-tarmsygdomme Nuværende lægemiddelmetabolisme, Bentham Science Publisher, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui, et al. Læk tarm som faresignal for autoimmune sygdomme Grænser, Frontiers, 5 Maj 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon, et al. Fermenterede fødevarer som en diætkilde til levende organismer Grænser i mikrobiologi, Frontiers Media SA, 24 Aug. 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., et al. Hurtig forstyrrelse af tarmbarrierefunktion ved Gliadin involverer ændret ekspression af apikale junctionelle proteiner. FEBS Press, John Wiley & Sons, Ltd, 8. august 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R Balfour. Terapeutisk manipulation af den enteriske mikroflora i inflammatoriske tarmsygdomme: antibiotika, probiotika og præbiotika. Gastroenterologisk, US National Library of Medicine, maj 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Kroniske smerter er et almindeligt sundhedsproblem, som påvirker mange mennesker i USA. Mens flere medicinske tilstande, såsom fibromyalgi og myofascial smertesyndrom, kan forårsage kroniske smerter, kan det også udvikle sig på grund af en række andre sundhedsproblemer. Forskningsundersøgelser har fundet ud af, at udbredt betændelse er den førende årsag til kroniske smerter. Betændelse er en naturlig forsvarsmekanisme mod skade, sygdom eller infektion. Men hvis den inflammatoriske proces fortsætter for længe, kan den blive problematisk.
Betændelse signalerer immunsystemet til at hele og reparere beskadiget væv samt at beskytte sig mod bakterier og vira. Som nævnt ovenfor kan kronisk betændelse dog forårsage en række sundhedsproblemer, herunder kroniske smertesymptomer. Sunde livsstilsændringer kan hjælpe med at håndtere kroniske smerter, men lad os først forstå de almindelige årsager til kronisk smerte.
Hvad er akut inflammation?
Akut betændelse opstår for eksempel efter en skade eller noget så simpelt som ondt i halsen. Det er en naturlig reaktion med negative virkninger, hvilket betyder, at den virker lokalt i den region, hvor sundhedsproblemet er fundet. De almindelige tegn på akut betændelse omfatter hævelse, rødme, varme, smerte og funktionstab, som angivet af National Library of Medicine. Når der udvikles akut betændelse, udvider blodkarrene sig, hvilket får blodgennemstrømningen til at øges, og hvide blodlegemer i den skadede region fremmer restitutionen.
Under alvorlig inflammation frigives forbindelser kaldet cytokiner af det beskadigede væv. Cytokinerne fungerer som "nødsignaler", som bringer menneskekroppens egne immunceller, såvel som hormoner og talrige næringsstoffer til at reparere sundhedsproblemet. Derudover forårsager hormonlignende stoffer, kendt som prostaglandiner, blodpropper til at hele beskadiget væv, og disse kan også udløse feber og smerter som en del af den inflammatoriske procedure. Efterhånden som skaden eller skaden kommer sig, aftager betændelsen.
Hvad er kronisk inflammation?
I modsætning til akut inflammation har kronisk inflammation langtidsvirkninger. Kronisk betændelse, også kendt som vedvarende inflammation, producerer lave niveauer af inflammation i hele menneskekroppen, som vist ved en stigning i immunsystemmarkører placeret i blod- og cellevæv. Kronisk betændelse kan også forårsage progression af forskellige sygdomme og tilstande. Forhøjede niveauer af inflammation kan nogle gange udløse, selvom der ikke er nogen skade, sygdom eller infektion, hvilket også kan få immunsystemet til at reagere.
Som et resultat kan den menneskelige krops immunsystem begynde at angribe sunde celler, væv eller organer. Forskere forsøger stadig at forstå konsekvenserne af kronisk inflammation i den menneskelige krop og de mekanismer, der er involveret i denne naturlige forsvarsproces. For eksempel er kronisk inflammation blevet forbundet med en række sundhedsproblemer, såsom hjertesygdomme og slagtilfælde.
En teori antyder, at når betændelse forbliver i blodkarrene, kan det tilskynde til ophobning af plak. Ifølge American Heart Association eller AHA, hvis immunsystemet identificerer plak som en fremmed angriber, kan de hvide blodlegemer forsøge at vægge af pladen, der findes i blodet, der strømmer gennem arterierne. Dette kan skabe en blodprop, som kan blokere blodtilførslen til hjertet eller hjernen, hvilket får det til at blive ustabilt og briste. Kræft er et andet sundhedsproblem forbundet med kronisk betændelse. Desuden kan DNA-skader ifølge National Cancer Institute også være forårsaget af kronisk inflammation.
Vedvarende, lavgradig inflammation har ofte ingen symptomer, men sundhedspersonale kan tjekke for et C-reaktivt protein eller CRP, kendt som lipoinsyre, en markør for betændelse fundet i blodet. Forhøjede niveauer af CRP er forbundet med en øget risiko for hjerte-kar-sygdomme. Forhøjede CRP-niveauer kan findes ved kroniske lidelser som lupus eller leddegigt.
I tilfælde af andre kroniske tilstande, såsom fibromyalgi, overreagerer nervesystemet på specifik stimulering, men det er betændelse, der forårsager kroniske smertesymptomer. Subjektivt er det næsten umuligt at kende forskel på den kroniske smerte forårsaget af et overfølsomt nervesystem og den kroniske smerte forårsaget af udbredt betændelse. Udover at søge efter spor i blodbanen, kan en persons ernæring, livsstilsvaner og miljøeksponeringer også fremme kronisk inflammation.
Betændelse er immunsystemets naturlige forsvarsmekanisme mod skader, sygdom eller infektion. Selvom denne inflammatoriske reaktion kan hjælpe med at helbrede og reparere væv, kan kronisk, udbredt betændelse forårsage en række sundhedsproblemer, herunder kroniske smertesymptomer. En afbalanceret ernæring, herunder en række forskellige diæter og faste, kan hjælpe med at reducere inflammation. Faste, også kendt som kaloriebegrænsning, fremmer celleapoptose og mitokondriel genopretning. Den fastende efterlignende diæt, som er en del af kostplanen for lang levetid, er et kostprogram, som "lurer" menneskekroppen til en fastende tilstand for at opleve fordelene ved traditionel faste. Før du følger nogen af de diæter, der er beskrevet i denne artikel, skal du sørge for at konsultere en læge.
Dr. Alex Jimenez DC, CCST Insight
Ernæring, diæter, faste og kroniske smerter
Anti-inflammatoriske diæter består hovedsageligt af at spise frisk frugt og grøntsager, fisk og fedt. Middelhavsdiætplanen er for eksempel en anti-inflammatorisk diæt, som fremmer spisning af moderate mængder nødder, indtagelse af meget lidt kød og vin. Anti-inflammatoriske fødevaredele, såsom omega-3 fedtsyrer, beskytter den menneskelige krop mod daMage forårsaget af betændelse.
En anti-inflammatorisk diæt indebærer også at holde sig væk fra fødevarer, der kan fremme betændelse. Det er ideelt at reducere mængden af fødevarer, du spiser, som er høj i trans og mættet fedt, såsom kød. Derudover begrænser en antiinflammatorisk diæt forbruget af raffinerede kulhydrater og fødevarer, såsom brød og ris. Disse fremmer også at skære ned på brugen af margarine og olier, der er fyldt med omega-6 fedtsyrer, såsom solsikke, tidsel. , majsolier.
Faste, eller kaloriebegrænsning, har længe været kendt for at mindske oxidativt stress og bremse mekanismerne for aldring i forskellige organismer. Effekterne af faste involverer programmeret celledød eller apoptose, transkription, mobil energieffektivitet, mitokondriel biogenese, antioxidantmekanismer og døgnrytme. Faste bidrager også til mitokondriel autofagi, kendt som mitofagi, hvor gener i mitokondrierne stimuleres til at gennemgå apoptose, hvilket fremmer mitokondriel genopretning.
Intermitterende faste kan hjælpe dig med at bekæmpe betændelse, forbedre fordøjelsen og øge din levetid. Den menneskelige krop er designet til at kunne overleve i længere tid uden mad. Forskningsundersøgelser har vist, at intermitterende faste kan have positive ændringer i den overordnede sammensætning af din tarmmikrobiota. Desuden kan intermitterende faste reducere insulinresistens og samtidig øge immunsystemets respons. Endelig kan intermitterende faste fremme produktionen af et stof, kendt som β-hydroxybutyrat, der blokerer en del af immunsystemet involveret i inflammatoriske lidelser samt væsentligt reducerer produktionen af inflammatoriske markører, såsom cytokiner og det C-reaktive protein , eller CRP, tidligere nævnt ovenfor.
The Longevity Diet Plan, præsenteret i bogen af Dr. Valter Longo, eliminerer forbruget af forarbejdede fødevarer, som kan forårsage betændelse, fremme velvære og lang levetid. Dette unikke kostprogram fremmer, i modsætning til de fleste traditionelle diæter, ikke vægttab. Selvom du kan opleve vægttab, ligger vægten af dette unikke kostprogram på at spise sundere. Longevity Diet Plan har vist sig at hjælpe med at aktivere stamcelle-baseret fornyelse, reducere abdominalt fedt og forhindre aldersrelateret knogle- og muskeltab, samt opbygge modstand mod udvikling af hjerte-kar-sygdomme, Alzheimers sygdom, diabetes og kræft.
Den fastende efterlignende diæt, eller MKS, giver dig mulighed for at opleve fordelene ved traditionel faste uden at fratage din krop mad. Den største forskel på mund- og klovsyge er, at i stedet for fuldstændig at eliminere al mad i flere dage eller endda uger, begrænser du kun dit kalorieindtag i fem dage om måneden. MKS kan praktiseres en gang om måneden for at fremme den generelle sundhed og velvære.
Mens alle kan følge MKS på egen hånd, ProLon fastende efterlignende diæt tilbyder et 5-dages måltidsprogram, som er blevet individuelt pakket og mærket for hver dag, som serverer de fødevarer, du har brug for til mund- og klovsyge i præcise mængder og kombinationer. Måltidsprogrammet består af klar til at spise eller let at tilberede, plantebaserede fødevarer, herunder barer, supper, snacks, kosttilskud, et drikkekoncentrat og te. Inden du starter ProLon� fastende efterlignende diæt, 5-dages måltidsprogram, eller nogen af de livsstilsændringer, der er beskrevet ovenfor, skal du sørge for at tale med en sundhedspersonale for at finde ud af, hvilken kronisk smertebehandling der passer til dig.
Omfanget af vores information er begrænset til kiropraktik, rygsøjlesundhedsproblemer og funktionel medicin artikler, emner og diskussioner. For yderligere at diskutere emnet ovenfor, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emnediskussion: Akut rygsmerter
Rygsmerte er en af de mest almindelige årsager til handicap og savnede dage på arbejdspladsen over hele verden. Rygsmerter tilskrives den næst mest almindelige årsag til lægebesøg, udelukkes kun ved øvre respiratoriske infektioner. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Din rygsøjle er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. Skader og / eller forværrede forhold, f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
XYMOGEN s Eksklusive Professional Formulas er tilgængelige via udvalgte licenserede sundhedspersonale. Internettet salg og diskontering af XYMOGEN formler er strengt forbudt.
stolt, Dr. Alexander Jimenez gør XYMOGEN formler kun tilgængelige for patienter under vores pleje.
Ring venligst til vores kontor for at give os en lægehøring om øjeblikkelig adgang.
Hvis du er patient af Skadesmedicinsk og kiropraktikklinik, kan du spørge om XYMOGEN ved at ringe 915-850-0900.
Til din bekvemmelighed og gennemgang af XYMOGEN produkter bedes du gennemse følgende link. *XYMOGEN-katalog-Hent
* Alle ovennævnte XYMOGEN-politikker forbliver strengt gældende.
Overholdelse af en bestemt diæt til opretholdelse af korrekt ernæring kan undertiden gøre spændingen stressende. Naturlige livsstilsændringer er nøglen til at ændre dine spisevaner, og dette kan hjælpe dig med at leve et længere og sundere liv. The Longevity Diet Plan, skabt af Dr. Valter Longo, er et udvalg af praktiske spise retningslinjer, der fokuserer på at ændre dine spisemønstre for at opnå overordnet sundhed og wellness.
Reglerne for livslang kostplan
Ved blot at følge de næringsmæssige tips nedenfor kan du revidere din nuværende diætplan og begynde at spise sundere uden al den stress i en traditionel kost. The Longevity Diet Plan eliminerer forbruget af forarbejdede fødevarer, der kan forårsage en række sundhedsproblemer og øger forbruget af næringsstoffer, der fremmer lang levetid. Dette unikke kostprogram deler resultaterne fra ca. 25 års forskningsundersøgelser alt sammen om en enkel løsning, der kan hjælpe folk med at opleve det generelle velfærd gennem korrekt ernæring.
I modsætning til de fleste traditionelle diæter fremmer Longevity Diet Plan imidlertid ikke vægttab. Selvom du måske oplever vægttab, lægges vægt på dette unikke diætprogram på at spise sundere. Longevity Diet Plan er blevet demonstreret for at hjælpe dig med at aktivere stamcellebaseret fornyelse, tabe sig og reducere mavefedt, forhindre aldersrelateret tab af knogler og muskler, opbygge modstand mod udvikling af hjerte-kar-sygdomme, Alzheimers sygdom, diabetes og kræft samt som forlænger levetiden. Nedenfor opsummerer vi de 8 mest almindelige ernæringstips i Longevity Diet Plan, som i sidste ende kan hjælpe med at gøre dit liv længere og sundere.
The Longevity Diet Plan er et unikt kostprogram designet af Dr. Valter Longo for at fremme overordnet sundhed, wellness og lang levetid. Gennem enkle livsstilsændringer kan folk ændre deres spisevaner og drage fordel af de mange sundhedsmæssige fordele ved dette diætprogram. Ved at følge en pescatarian diæt og følge ProLon Fastende efterlignende diæt, blandt de andre næringsmæssige tips, der er beskrevet nedenfor, kan folk leve længere og sundere liv. Traditionelle kostvaner kan ofte være vanskelige og stressfulde at følge, men Longevity Diet Plan er et praktisk og unikt kostprogram, som kan være velegnet til mange mennesker.
Dr. Alex Jimenez DC, CCST Insight
8 Ernæringsmæssige Tips til Livsplanen
Følg en Pescatarian Diet
Som en del af lang levetid planen, følg en pescatarian diæt, som er næsten 100 procent plante og fisk-baseret. Sørg også for at begrænse fiskforbruget til to eller tre portioner hver uge, så du undgår fisk med højere indhold af kviksølv, som tun, sværdfisk, makrel og hellefisk. Hvis du er over 65, og du begynder at opleve nedsat muskelmasse, styrke og fedt, skal du tilføje flere fisk i din kost sammen med andre dyrebaserede fødevarer, herunder æg og bestemte oste, såsom feta eller pecorino og yoghurt fremstillet af gedens mælk.
Spis ikke for meget protein
Ifølge Longevity Diet Plan bør vi spise 0.31 til 0.36 gram protein pr. Pund kropsfedt hver dag. Hvis du vejer 130lbs, skal du spise omkring 40 til 47 gram protein pr dag, eller et tilsvarende 1.5 fileter af laks, 1 kop kikærter eller 2 1 / 2 kopper linser, hvoraf 30 gram skal indtages i et måltid. Hvis du vejer 200 til 220lbs, skal du spise omkring 60 til 70 gram protein pr. Dag eller et tilsvarende af to laksfilet, 3 1 / 2 kopper linser eller 1 1 / 2 kopper kikærter. Proteinforbruget bør øges efter alder 65. For de fleste af os er en 10 til 20 procent stigning, eller 5 til 10 gram mere hver dag, nok. Endelig er lang levetid kostfri med animalske proteiner som rødt kød, hvidt kød og fjerkræ med undtagelse af animalske proteiner i fisk. Dette unikke kostprogram er i stedet forholdsvis højt i vegetabilske proteiner som belgfrugter og nødder for at optimere sundhed og wellness.
Forøg god fedt og komplekse kulhydrater
Som en del af langtidsplejeplanen bør du spise større mængder af flerumættede fedtstoffer, som dem, der findes i laks, mandler, valnødder og olivenolie, mens du skal spise lavere mængder af mættede, hydrogenerede og transfedtstoffer. Ligeledes bør du som en del af langtidsplejeplanen spise komplekse kulhydrater, som dem, der findes i fuld hvedebrød, bælgfrugter og grøntsager. Sørg for at begrænse spiser pasta, ris, brød, frugt og frugtsaft, som kan omdannes til sukkerarter, når de når din tarme.
Tag kosttilskud
Den menneskelige krop har brug for proteiner, essentielle fedtsyrer som omega-3 og omega-6, vitaminer, mineraler og endog sukkerarter til at fungere korrekt. Når dit indtag af visse næringsstoffer bliver for lavt, kan reparations-, udskiftnings- og forsvarsmetoderne i den menneskelige krop sænke eller stoppe, så svampe, bakterier og vira kan forårsage skade, der kan medføre en række sundhedsmæssige problemer. Tag vitamin og mineral kosttilskud, især til omega-3, som anbefalet af din sundhedspersonale.
Spis forskellige fødevarer fra din Ancestry
At tage i alle de nødvendige næringsstoffer, du har brug for, skal du spise en bred vifte af fødevarer, men det er bedst at vælge fødevarer, der var almindelige på din forældres, bedsteforældres og bedsteforældres bord. I mange nordøsteuropæiske lande, hvor mælk generelt er blevet brugt, er lactoseintolerans relativt sjælden, mens lactoseintolerans er ret almindelig i sydeuropæiske og asiatiske lande, hvor mælk ikke historisk var en del af den konventionelle kost af voksne. Hvis en person med japansk forfædre bosiddende i USA pludselig beslutter sig for at begynde at drikke mælk, som sandsynligvis sjældent blev betjent i deres bedsteforældres spisebord, vil de sandsynligvis begynde at blive syge. De mest almindelige problemer i disse tilfælde er intolerancer eller autoimmuniteter, såsom reaktionen på glutenrige fødevarer som brød og pasta set hos mennesker med celiaciasygdom. Selv om der er brug for yderligere beviser, er det muligt, at fødeintolerancer kan være relateret til mange autoimmune sygdomme, herunder diabetes, colitis og Crohns sygdom.
Spis to måltider om dagen og en snack
I henhold til Longevity Diet Plan er det ideelt at spise morgenmad og et større måltid plus en nærende snack med lavt kalorieindhold og lavt sukkerindhold hver dag. Mens det for nogle mennesker kan anbefales at spise tre måltider og en snack hver dag. Mange ernæringsretningslinjer anbefaler, at vi spiser fem til seks måltider hver dag. Når folk rådes til at spise ofte, kan det ofte blive svært for dem at regulere deres kalorieindtag. I løbet af de sidste tyve år anses ca. 70 procent af befolkningen i USA for at være overvægtige eller overvægtige. Det er meget sværere at overspise på Longevity Diet Plan, hvis du kun spiser to og et halvt måltid hver dag. Det ville tage massive portioner bælgfrugter, grøntsager og fisk at nå det beløb, der ville føre til vægtøgning. Den høje næring af måltiderne plus mængden af måltidet sender et signal til din mave og din hjerne om, at du har fået nok mad. Dette ene store måltidsystem kan undertiden blive opdelt i to måltider for at undgå fordøjelsesproblemer. Voksne og ældre, der er tilbøjelige til vægttab, bør spise tre måltider om dagen. For folk, der prøver at tabe sig såvel som for folk, der er overvægtige eller overvægtige, er det bedste ernæringsråd at spise morgenmad dagligt; spis middag eller frokost, men ikke begge dele, og erstat det ubesvarede måltid med en snack, der indeholder færre end 100 kalorier og ikke mere end 3 til 5 g sukker. Hvilket måltid du springer over afhænger af din livsstil, men det anbefales ikke at springe morgenmaden over på grund af dens negative sundhedsmæssige problemer. Fordelen ved at springe over frokost er mere fritid og energi. Men der er en ulempe ved at spise en stor middag, især for folk, der lider af syre refluks eller søvnproblemer. Ulempen ved at springe over middagen er imidlertid, at det kan eliminere det sociale måltid på deres dag.
Spis inden for et 12-timers vindue hver dag
En anden almindelig spisestue vedtaget af mange centenarer er tidsbegrænset spiser eller begrænser alle måltider og snacks inden for et 12-timers vindue hver dag. Effektiviteten af denne metode blev påvist i både undersøgelser af mennesker og dyr. Generelt ville du spise morgenmad på 8 am og derefter spise middag ved 8 pm. Et briefer spise vindue på ti timer eller mindre kan være endnu bedre for vægttab, men det er betydeligt sværere at vedligeholde, og det kan øge risikoen for at udvikle bivirkninger, såsom gallesten og endda potentielt øge chancen for at udvikle hjerte-kar-sygdomme. Du bør ikke spise tre til fire timer før du sover.
Følg ProLon Fastefterlignende diæt
Friske mennesker under 65 'alder skal følge ProLon Fastende efterlignende diæt, 5-dages måltidsprogram mindst to gange om året. MHD er et af de nøgleprincipper, der fremmes af langtidsplejeplanen. Den fastende efterligne diæt giver de samme sundhedsmæssige fordele ved at faste uden at faste. Ved at spise 800 til 1,100 kalorier i præcise mængder og kombinationer af fødevarer, der er pakket individuelt og mærket for hver dag, kan du "trick" menneskekroppen i en fastende tilstand. Gennem forskellige forskningsundersøgelser har Dr. Valter Longo opdaget, at ved at berøve kroppen af mad på denne måde, begynder vores celler at nedbryde og regenerere vores indre væv gennem en proces, der kaldes autofagi, dræbe og erstatte eller regenerere beskadigede celler. Derudover kan fasting reversere forskellige sundhedsmæssige problemer, ødelægge kræftceller og reducere muligheden for at udvikle Alzheimers sygdom betydeligt.
Med Longevity Diet Plan præsenteret i bogen af Dr. Valter Longo spiser du bedre, føler dig bedre, og selvom den ikke er designet som et vægttabsplan, kan du endda kaste et par pund. Du bliver ikke nødt til at overveje komplekse madregler og træffe vanskelige valg med dette unikke diætprogram. Når du først har fået fat på disse livsstilsændringer, kan du forbedre din generelle sundhed og velvære såvel som din levealderen. omfanget af vores information er begrænset til kiropraktik, spinal sundhed spørgsmål og funktionelle medicin emner. For yderligere at diskutere emnet, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emnediskussion: Akut rygsmerter
Rygsmerte er en af de mest almindelige årsager til handicap og savnede dage på arbejdspladsen over hele verden. Rygsmerter tilskrives den næst mest almindelige årsag til lægebesøg, udelukkes kun ved øvre respiratoriske infektioner. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Din rygsøjle er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. Skader og / eller forværrede forhold, f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
XYMOGEN s Eksklusive Professional Formulas er tilgængelige via udvalgte licenserede sundhedspersonale. Internettet salg og diskontering af XYMOGEN formler er strengt forbudt.
stolt, Dr. Alexander Jimenez gør XYMOGEN formler kun tilgængelige for patienter under vores pleje.
Ring venligst til vores kontor for at give os en lægehøring om øjeblikkelig adgang.
Hvis du er patient af Skadesmedicinsk og kiropraktikklinik, kan du spørge om XYMOGEN ved at ringe 915-850-0900.
Til din bekvemmelighed og gennemgang af XYMOGEN produkter bedes du gennemse følgende link. *XYMOGEN-katalog-Hent
* Alle ovennævnte XYMOGEN-politikker forbliver strengt gældende.
IFM's Find A Practitioner-værktøj er det største henvisningsnetværk inden for Functional Medicine, skabt for at hjælpe patienter med at finde Functional Medicine-praktikere overalt i verden. IFM Certified Practitioners er anført først i søgeresultaterne på grund af deres omfattende uddannelse i Functional Medicine