
Vi blev alle undervist som børn, at der er 5 sanser: syn, smag, lyd, lugt og berøring. De første fire sanser bruger klare, adskilte organer, som f.eks. Øjne, smagsløg, ører og næse, men hvordan virker kropssansen nøjagtigt? Touch er oplevet over hele kroppen, både indenfor og udenfor. Der er ikke et særskilt organ, der er ansvarlig for at føle berøring. Der er snarere små receptorer eller nerveender rundt omkring i hele kroppen, som fornemmer at røre ved hvor det forekommer og sender signaler til hjernen med oplysninger om den type berøring der opstod. Som smagsprøve på tungen opdager smag er mekanoreceptorer kirtler i huden og på andre organer, der registrerer følelser af berøring. De er kendt som mekanoreceptorer fordi de er designet til at opdage mekaniske fornemmelser eller forskelle i tryk.
Indhold
Rolle af mekanoreceptorer
En person forstår, at de har oplevet en fornemmelse, når organet der er ansvarlig for at opdage, at den specifikke forstand sender en besked til hjernen, som er det primære organ, som behandler og arrangerer al information. Meddelelser sendes fra alle områder af kroppen til hjernen gennem ledninger benævnt neuroner. Der er tusindvis af små neuroner, der forgrener sig til alle områder af menneskekroppen, og på slutningen af mange af disse neuroner er mekanoreceptorer. For at demonstrere, hvad der sker, når du rører et objekt, bruger vi et eksempel.
Forestil dig en mygeland på din arm. Stammen af dette insekt, så let, stimulerer mekanoreceptorer i det pågældende område af armen. Disse mekanoreceptorer sender en besked langs den neuron, de er forbundet med. Neuronet forbinder hele vejen til hjernen, som modtager meddelelsen om, at noget berører din krop i den nøjagtige placering af den specifikke mekanoreceptor, der sendte beskeden. Hjernen vil handle med dette råd. Måske vil det fortælle øjnene at se på den region af armen, der registrerede signaturen. Og når øjnene fortæller hjernen, at der er en myg på armen, kan hjernen fortælle hånden om hurtigt at smide det væk. Sådan virker mekanoreceptorer. Formålet med artiklen nedenfor er at demonstrere og diskutere i detaljer den funktionelle organisering og molekylære determinanter for mekanoreceptorer.
Touch Sense: Funktionel organisation og molekylære determinanter af mekanosensitive receptorer
Abstrakt
Kutane mekanoreceptorer er lokaliseret i de forskellige lag i huden, hvor de opdager en bred vifte af mekaniske stimuli, herunder lys børste, stræk, vibration og skadeligt tryk. Denne variation af stimuli matches af et mangfoldigt udvalg af specialiserede mekanoreceptorer, der reagerer på kutan deformation på en bestemt måde og relæer disse stimuli til højere hjernestrukturer. Undersøgelser på tværs af mekanoreceptorer og genetisk indgreb i sensoriske nerveender begynder at afdække touch sensationsmekanismer. Arbejdet på dette område har givet forskere en mere grundig forståelse af kredsløbsorganisationen, der ligger til grund for opfattelsen af berøring. Nye ionkanaler er opstået som kandidater til transduktionsmolekyler og egenskaber af mekanisk gatedstrømme forbedrede vores forståelse af mekanismerne til tilpasning til taktile stimuli. Denne gennemgang fremhæver de fremskridt, der er gjort med at karakterisere funktionelle egenskaber hos mekanoreceptorer i hårede og glabro hud- og ionkanaler, der registrerer mekaniske indgange og form mekanoreceptor tilpasning.
nøgleord: mekanoreceptor, mekanosensitiv kanal, smerte, hud, somatosensorisk system, touch
Introduktion
Touch er påvisning af mekanisk stimulering, der påvirker huden, herunder uskadelige og skadelige mekaniske stimuli. Det er en væsentlig fornuft for overlevelse og udvikling af pattedyr og mennesker. Kontakt med faste genstande og væsker med huden giver nødvendig information til centralnervesystemet, der muliggør efterforskning og anerkendelse af miljøet og initierer bevægelse eller planlagt håndbevægelse. Touch er også meget vigtigt for lærlingeforhold, sociale kontakter og seksualitet. Berøringsfølsomhed er den mindst sårbare fornemmelse, selv om den kan forvrænges (hyperesthesi, hypoestesi) under mange patologiske forhold. 1-3
Berøringssvar involverer en meget præcis kodning af mekanisk information. Kutane mekanoreceptorer er lokaliseret i de forskellige lag af huden, hvor de registrerer en bred vifte af mekaniske stimuli, herunder let børste, strækning, vibration, afbøjning af hår og skadeligt tryk. Denne række stimuli matches af en række forskellige specialiserede mekanoreceptorer, der reagerer på kutan deformation på en bestemt måde og videreformidler disse stimuli til højere hjernestrukturer. Somatosensoriske neuroner i huden falder i to grupper: mekanismer med lav tærskel (LTMR'er), der reagerer på godartet tryk, og højterskel mekanoreceptorer (HTMR'er), der reagerer på skadelig mekanisk stimulering. LTMR- og HTMR-cellekroppe befinder sig i dorsal rodganglier (DRG) og kraniale sensoriske ganglier (trigeminusganglier). Nervefibre associeret med LTMR'er og HTMR'er klassificeres som Aa -, Aa - eller C-fibre baseret på deres handlingspotentiale ledningshastigheder. C-fibre er umyeliniserede og har de langsomste ledningshastigheder (~ 2 m / s), mens A? og A? fibre er let og stærkt myeliniserede og udviser henholdsvis mellemliggende (~ 12 m / s) og hurtige (~ 20 m / s) ledningshastigheder. LTMR'er klassificeres også som langsomt eller hurtigt tilpasningsresponser (SA- og RA-LTMR'er) i henhold til deres tilpasningshastigheder til vedvarende mekanisk stimulus. De adskiller sig yderligere ved de kutane endeorganer, de innerverer, og deres foretrukne stimuli.
Mulighed for mekanoreceptorer til at detektere mekaniske signaler er afhængig af tilstedeværelsen af mekanotransducer ionkanaler, der hurtigt omdanner mekaniske kræfter til elektriske signaler og depolariserer det modtagelige felt. Denne lokale depolarisation, der kaldes receptorpotentiale, kan generere actionpotentialer, der formerer sig mod centralnervesystemet. Egenskaber af molekyler, der formidler mekanotransduktion og tilpasning til mekaniske kræfter forbliver uklare.
I denne anmeldelse giver vi en oversigt over pattedyrs mekanoreceptoregenskaber i uskadelig og skadelig berøring i den hårede og glat hud. Vi overvejer også den nylige viden om egenskaberne ved mekanisk styrede strømme i et forsøg på at forklare mekanismen til mekanoreceptors tilpasning. Endelig gennemgår vi de seneste fremskridt med at identificere ionkanaler og tilknyttede proteiner, der er ansvarlige for genereringen af mechano-gatede strømme.
Innocuous Touch
Hårfollikel-associerede LTMR'er
Hårsækkene repræsenterer hårsaksproducerende mini-organer, der opdager let berøring. Fibre, der er forbundet med hårsækkene, reagerer på hårbevægelse og dets retning ved at skyde tog af handlingspotentialer ved begyndelsen og fjernelsen af stimulus. De tilpasser hurtigt receptorer.
Katte og kanin. I katte- og kaninfrakke kan hårsækkene opdeles i tre hårsækketyper, Down-hår, Guard-hår og Tylotrichs. Nedhårene (underhår, uld, vellus) 4 er de mest talrige, de korteste og fineste hår i pelsen. De er bølgede, farveløse og fremkom i grupper på to til fire hår fra en almindelig åbning i huden. Vagthårene (monotrichs, overhør, tophair) 4 er let buede, enten pigmenterede eller upigmenterede og kom enkeltvis ud af munden på deres follikler. Tylotrichs er de mindst talrige, de længste og tykkeste hår.5,6 De er pigmenterede eller upigmenterede, nogle gange begge og kom enkeltvis ud af en follikel, der er omgivet af en løkke med kapillære blodkar. De sensoriske fibre, der leveres til en hårsække, er placeret under talgkirtlen og tilskrives A? eller A? -LTMR fibre.7
I umiddelbar nærhed til den nedre håraksel er lige under niveauet af talgkirtlen ringet af lancetformede pilo-Ruffini-slutninger. Disse sensoriske nerveender er anbragt i en spiralbane rundt om hårets aksel i bindevævet, der danner hårsækken. Inden for hårsækken er der også frie nerveender, hvoraf nogle danner mekanoreceptorer. Hyppigt, berør corpuscles (se glabro hud) omkring halsområdet af tylotrich follikel.
Egenskaber for myeliniserede nerveender i hår af kat og kanin er blevet udforsket intenst i perioden 1930-1970 (gennemgang i Hamann, 1995). 8 Bemærkelsesværdigt, Brown og Iggo, der studerede 772 enheder med myeliniserede afferente nervefibre i de saphenøse nerver fra kat og kanin, har klassificeret respons i tre receptortyper svarende til bevægelserne af Down-hår (type D-receptorer), Guard-hår (type G-receptorer) og Tylotrich-hår (type T-receptor) .9 Alle de afferente nervefibersvar er blevet samlet i den hurtigt tilpasningsreceptor af type I (RA I) ved modstand mod den Pacinian-receptor ved navn RA II. RA I mekanoreceptorer registrerer hastigheden af mekanisk stimulus og har skarp kant. De registrerer ikke termiske variationer. Burgess et al. beskrev også en hurtigt tilpasende feltreceptor, der reagerer optimalt på strøgning af huden eller bevægelse af flere hår, hvilket blev tilskrevet stimulering af pilo-Ruffini-ender. Intet af hårsækkets respons blev tilskrevet C-fiberaktivitet.10
Mus. I musens dorsale hårde hud er tre hovedtyper af hårsække blevet beskrevet: zigzag (omkring 72%), awl / auchene (omkring 23%) og vagt eller tylotrich (omkring 5%). 11-14 Zigzag og Awl / auchenne hårsække producerer de tyndere og kortere håraksler og er forbundet med en talgkirtlen. Guard eller tylotrich hår er den længste af hårsække typer. De er præget af en stor hårpære forbundet med to talgkirtler. Vagt og A / S hår er arrangeret i et iterativt, regelmæssigt mellemrummet mønster, mens zigzaghår tætbefolket hudområder omkring de to større hårfollikeltyper [Fig. 1 (A1, A2 og A3)].
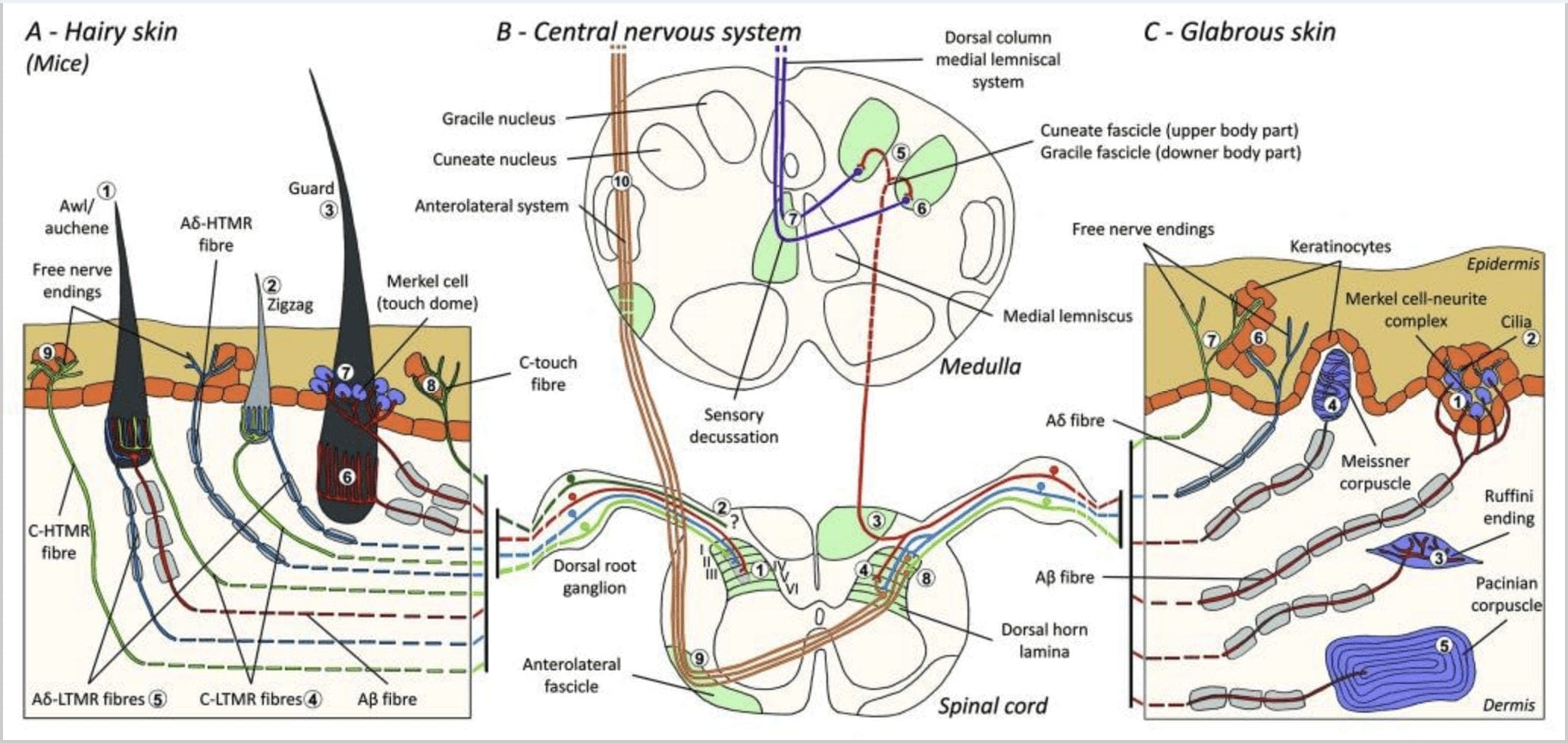
Figur 1. Organisation og fremskrivninger af kutane mekanoreceptorer. I hårig hud opdages lyspensel og berøring hovedsageligt af innerveringen omkring hårsækkene: awl / auchenne (A1), zigzag (A2) og vagt (A3). Awl / auchene-hår er tredobbelt innerveret af C-LTMR-lancetformede ender (A4), A? -LTMR og A? hurtigt tilpasning-LTMR (A6). Zigzag-hårsækkene er de kortere håraksler og er innerveret af både C-LTMR (A4) og A? -LTMR lancetformede ender (A5). De længste vagt hårsækkene er innerveret af A? hurtigt tilpasning-LTMR langsgående lancetformede ender (A6) og er forbundet med A? langsomt tilpasning-LTMR af touch dome-ender (A7). De centrale fremspring af alle disse fibre slutter i distinkte, men delvist overlappende laminater i rygmarvets dorsale horn (C-LTMR i lamina II, Aa-LTMR i lamina III og Aa-LTMR i lamina IV og V). Fremspringene af LTMR, som innerverer de samme eller tilstødende hårsækkede stoffer, er justeret for at danne en smal søjle i rygmarvets dorsale horn (B1 i grå). Kun i hårig hud indvander en subpopulation af C-fibre frie ende epidermis og reagerer på behagelige berøring (A8). Disse C-touch-fibre reagerer ikke på skadelig berøring, og deres vejkørsel er endnu ikke kendt (B2). I glabro hud medieres uskadelig berøring af fire typer LTMR'er. Merkel-celle-neurit-komplekset er i det basale lag af epidermis (C1). Denne mekanoreceptor består af et arrangement mellem mange Merkel-celler og en forstørret nerveterminal fra en enkelt A? fiber. Merkelceller udviser fingerlignende processer, der kontakter keratinocytter (C2). Ruffini-slutningen er lokaliseret i dermis. Det er en tynd cigarformet indkapslet sensorisk slutning forbundet med A? fiber (C3). Meissner-legemet forbundet med A? nerveender og er placeret i dermale papiller. Denne indkapslede mekanoreceptor består af pakket ned understøttende celler indrettet som vandrette lameller omgivet af bindevæv (C4). Pacinian corpuscle er den dybere mekanoreceptor. En enkelt A? umyelineret nerveende slutter i midten af denne store ovoide blodlegeme lavet af koncentriske lameller. Fremskrivninger af disse A? -LTMR-fibre i rygmarven er opdelt i to grene. Den vigtigste centrale gren (B3) stiger i rygmarven i den ipsilaterale dorsale danner cuneate eller gracile hæfterne (B5) upon medulla niveau, hvor de primære afferenter gør deres første synapse (B6). De sekundære neuroner gør en sensorisk decussation (B7) til dannelse af en kanal på den mediale lemniscus, der stiger gennem hjernestammen til midterbenet, specielt i thalamus. Den sekundære branche af LTMR ophører i dorsalhornet i lamina II, IV, V og forstyrrer smertetransmissionen (B4). Kræftende berøring opdages af den frie nerve ende i epidermis af begge hårede (A9) og glabro hud (C7). Disse mekanoreceptorer er afslutningen på Aa-HTMR og C-HTMR i tæt kontakt med nærliggende keratinocytter (C6). Aa-hTMR slutter i lamellen I og V; C-HTMR afsluttes i lamina I og II (B8). På rygmarvsdorsalhornsniveau producerer primære afferenter HTMR synapser med sekundære neuroner, som krydser midterlinien og klatre op til den højere hjernestruktur i den anterolaterale fascikel (B9, B10).
For nylig brugte Ginty og samarbejdspartnere en kombination af molekylær-genetisk mærkning og somatotopiske retrograd sporingsmetoder til at visualisere organisationen af perifere og centrale aksonale ender af LTMR'erne hos mus.15 Deres fund understøtter en model, hvor individuelle træk ved en kompleks taktil stimulus er ekstraheret af de tre hårsækketyper og transporteret via aktiviteterne i unikke kombinationer af A? -, A? - og C-fibre til dorsalt horn.
De viste, at den genetiske mærkning af tyrosinhydroxylase-positive (TH +) DRG-neuroner karakteriserer en population af ikke-peptidergiske sensoriske neuroner med lille diameter og giver mulighed for visualisering af C-LTMR perifere ender i huden. Overraskende viste det sig, at de axoneale grene af individuelle C-LTMR'er arboriserede og dannede langsgående lansformede ender, der er tæt forbundet med zigzag (80% af enderne) og syl / auchen (20% af enderne), men ikke tylotrich hårsækkene [Fig. 1 (A4)]. Langsgående lansetformede ender har længe været anset for at høre udelukkende til A? -LTMR'er, og det var derfor uventet, at enderne af C-LTMR'er ville danne langsgående, lansettede ender.15 Disse C-LTMR'er har en mellemliggende tilpasning i sammenligning med de langsomt og hurtigt tilpasning myeliniserede mekanoreceptorer [fig. 2 (C1)].
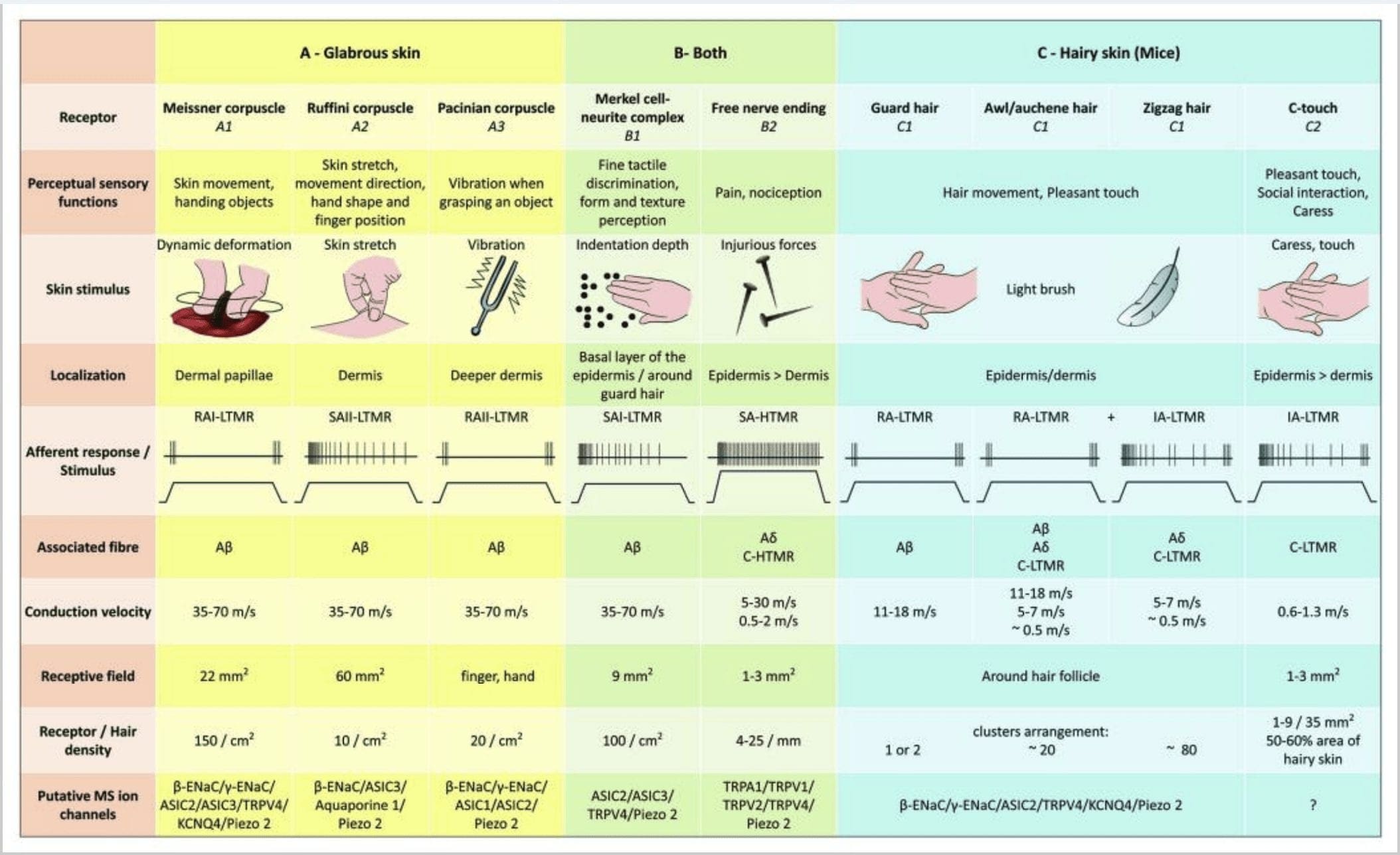
Figur 2. Taktile receptorer i pattedyr: Kutane taktile receptorer adskiller sig til uskadelig berøring understøttet af flere receptorer med lav mekanisk tærskel (LTMR'er) i glat og håret hud og skadelig berøring understøttet af højmekanisk tærskelreceptor (HTMR'er). De udgør nervefri ender, der primært ender i epidermis. (A) Glat hud. A1: Meissner-legemer registrerer hudbevægelse og glider af genstand i hånden. De er vigtige for at aflevere objekt og fingerfærdighed. Receptorer tilpasser sig hurtigt til stimulus, er forbundet med A? fibre og tyndt til C-fibre og har stort receptorfelt. A2: Ruffini-legemer registrerer strækning af huden og er vigtige for at opdage fingerposition og afleveringsgenstand. Receptoren tilpasser sig langsomt til stimulus og opretholdt aktivitet, så længe stimulus blev anvendt. Receptorer er forbundet til A? fibre og har stort modtageligt felt. A3: Paciniske legemer er dybere i dermis og registrerer vibrationer. Receptorer er forbundet til A? fibre; de tilpasser sig hurtigt til stimulus og har det største modtagelige felt. (B) Hele huden. B1: Merkel-cellekomplekser er til stede i både glat hud og omkring hår. De er tæt udtrykt i hånden og er vigtige for teksturopfattelse og fineste diskrimination mellem to punkter. De er ansvarlige for fingerpræcision. Receptorer er forbundet til A? fibre; de tilpasser sig langsomt til stimulus og har et kort modtageligt felt. B2: skadelige berørings-HTMR'er med meget langsom tilpasning til stimulus, dvs. aktive, så længe den nociceptive stimulus anvendes. De er dannet af den frie nerveende af A? og C-fibre associeret med keratinocytter. (C) Behåret hud. C1: Hårsækkene er forbundet med de forskellige hårtyper. Hos mus er beskyttelseshår den længste og tyndt udtrykte, syl / auchenne er af mellemstørrelse, og zigzag er det mindste og tættest udtrykte hår. De er forbundet til A? fibre men også til A? og C-LTMRs fibre til syl / auchenne og zizag hår. De registrerer hårbevægelser inklusive behagelig berøring under kærtegn. De tilpasser sig hurtigt eller med mellemliggende kinetiske til stimulus. C2: C-touch nerveender svarer til en undertype af C-fibre terminal med fri slutning karakteriseret ved en lav mekanisk tærskel. De formodes at kode for behagelig fornemmelse fremkaldt af kærtegn. De tilpasser sig moderat til stimulus og har et kort modtageligt felt. Formodede mekanosensitive (MS) ionkanaler udtrykt i de forskellige taktile receptorer er angivet i overensstemmelse hermed til foreløbige data og opsummerer den nuværende hypotese under evaluering.
En anden identificeret større population vedrører A? -LTMR-enderne i Awl / Auchenne og zigzag-follikler, der skal sammenlignes med Down-hårsækken, som er grundigt undersøgt hos kat og kanin. Ginty og samarbejdspartnere viste, at TrkB udtrykkes ved høje niveauer i en delmængde af medium-diametre DRG-neuroner. Intracellulære optagelser ved hjælp af ex vivo hud-nervepræparation af mærkede fibre afslørede, at de udviser de fysiologiske egenskaber af fibre, der tidligere er undersøgt hos kat og kanin: udsøgt mekanisk følsomhed (Von Frey-tærskel <0.07 mN), der hurtigt tilpasser responser til suprathreshold stimuli, mellemledning hastigheder (5.8 0.9 m / s) og smalle ikke-bøjede soma pigge.15 Disse A? -LTMR'er danner langsgående lansetformede ender forbundet med stort set alle zigzag og syl / auchen hårsækken i bagagerummet [fig. 1 (A5)].
Endelig viste de, at de perifere ender ved hurtigt at tilpasse A? LTMR'er danner langsgående lansformede ender forbundet med vagt (eller tylotrich) og syl / auchen-hårsækkene [fig. 1 (A6)]. 15 Derudover er vagthår også forbundet med et Merkel-cellekompleks, der danner en berøringshoved forbundet med A? langsomt tilpasning af LTMR [fig. 1 (A7)].
Sammenfattende er næsten alle zigzag-hårsækkene innerveret af både C-LTMR- og Aa-LTMR-lancetformede ender; awl / auchene hår er tredobbelt innerveret af A? hurtig tilpasning-LTMR, Aa-LTMR og C-LTMR lancetformede ender; Guard hårsækkene er innerveret af A? hurtigt tilpasning-LTMR langsgående lansetformede ender og interagere med A? langsomt tilpasning-LTMR af touch dome-ender. Således modtager hver musefollikel unikke og uforanderlige kombinationer af LTMR-ender svarende til neurofysiologisk adskilte mekanosensoriske endeorganer. I betragtning af det iterative arrangement af disse tre hårtyper foreslår Ginty og samarbejdspartnere, at behåret hud består af iterativ gentagelse af perifer enhed, der indeholder, (1) et eller to centralt beliggende beskyttelseshår, (2) ~ 20 omgivende syl / auchennehår og (3 ) ~ 80 islagede zigzaghår [fig. 2 (C1)].
Rygmarv fremspring. De centrale fremskrivninger af A? hurtigt tilpasnings-LTMR'er, Aa-LTMR'er og C-LTMR'er afsluttes i distinkte, men delvist overlappende laminater (II, III, IV) i rygmarvets dorsale horn. Derudover er de centrale terminaler af LTMR'er, der innerverer de samme eller tilstødende hårsækkene i en perifer LTMR-enhed, justeret for at danne en smal LTMR-søjle i rygmarvets dorsale horn [fig. 1 (B1)]. Det ser således ud til, at en kile eller søjle af somatotopisk organiserede primære sensoriske afferente ender i dorsalhornet repræsenterer justeringen af de centrale fremspring af A? -, A? - og C-LTMR'er, der innerverer den samme perifere enhed og detekterer mekanisk stimuli, der virker på den samme lille gruppe af hårsækkene. Baseret på antallet af beskyttelses-, syl / auchen- og zigzaghår i bagagerummet og lemmerne og antallet af hver LTMR-undertype, anslår Ginty og samarbejdspartnere, at musens dorsale horn indeholder 2,000 4,000 LTMR-søjler, hvilket svarer til det omtrentlige antal perifere enheder LTMR-enheder.15
Endvidere er axoner af LTMR-undertyper tæt forbundet med hinanden med sammenflettede fremspring og interdigiterede lansetformede ender, der innerverer den samme hårsækken. Derudover, fordi de tre hårfollikeltyper udviser forskellige former, størrelser og cellulære sammensætninger, vil de sandsynligvis have tydelige afbøjnings- eller vibrationsindstillingsegenskaber. Disse fund er i overensstemmelse med klassiske neurofysiologiske målinger hos katten og kaninen, hvilket indikerer, at A? RA-LTMR'er og Aa-LTMR'er kan aktiveres differentielt ved afbøjning af forskellige hårsækketyper.16,17
Afslutningsvis er touch i håret hud kombinationen af: (1) de relative tal, unikke rumlige udbredelser og forskellige morfologiske og deflekterende egenskaber af de tre typer hårsække; (2) de unikke kombinationer af LTMR subtype-slutninger associeret med hver af de tre hårfollikeltyper; og (3) særskilte følsomheder, ledningshastigheder, spike-togmønstre og tilpasningsegenskaber af de fire hovedklasser af hårfollikelassocierede LTMR'er, som gør det hårde hudmekanosensory-system ekstraheret og overført til CNS de komplekse kombinationer af kvaliteter, som definerer en røre ved.
Free-Nerve Endings LTMRs
Generelt er C-fibre frie ender i huden HTMR'er, men en underpopulation af C-fibre reagerer ikke på skadelig berøring. Denne delmængde af taktile C-fibre (CT) afferenter repræsenterer en særskilt type umyeliniserede, lavterskel mekanoreceptive enheder, der findes i den hårede men ikke glatte hud hos mennesker og pattedyr [fig. 1 (A8)]. 18,19 CT'er er generelt forbundet med opfattelsen af behagelig taktil stimulering i kropskontakt.20,21
CT-afferenter reagerer på fordybningskræfter i området 0.3 2.5 mN og er således lige så følsomme over for huddeformation som mange af A? afferenter.19 Tilpasningskarakteristikaene for CT-afferenter er således mellemliggende i sammenligning med de langsomt og hurtigt tilpasning af myeliniserede mekanoreceptorer. De receptive felter af humane CT-afferenter er omtrent runde eller ovale i form. Feltet består af en til ni små responsive pletter fordelt over et område op til 35 mm2.22 Mushomologreceptorerne er organiseret i et mønster af diskontinuerlige pletter, der dækker ca. 50% af arealet i den hårede hud [fig. 60 (C2)]. 2
Bevis fra patienter, der mangler myelinerede taktile afferenter, indikerer, at signalering i CT-fibre aktiverer den økologiske cortex. Da dette system er dårligt ved kodning af diskriminerende aspekter ved berøring, men velegnet til kodning af langsomt og forsigtigt berøring, kan CT-fibre i hårig hud være en del af et system til behandling af behagelige og samfundsmæssige relevante aspekter af berøring. 24 CT-fiberaktivering kan også har en rolle i smertehæmning, og det er for nylig blevet foreslået, at betændelse eller traume kan ændre sensationen transporteret af C-fiber LTMR'er fra behagelig berøring til smerte. 25,26
Hvilken pathway CT-afferents rejse er endnu ikke kendt [Fig. 1 (B2)], men lavt tærskel-taktile indgange til spinothalamiske projektionsceller er blevet dokumenteret. 27-udlån anerkender rapporter om subtile kontralaterale underskud af trykdetektering hos menneskelige patienter efter ødelæggelse af disse veje efter chordotomi-procedurer. 28
LTMR i glabro hud
Merkel celle-neurit komplekser og touch dome. Merkel (1875) var den første til at give en histologisk beskrivelse af klynger af epidermale celler med store lobulerede kerner, der kom i kontakt med formodede afferente nervefibre. Han antog, at de underkendte følelse af berøring ved at kalde dem Tastzellen (taktile celler). Hos mennesker beriges Merkel-celleneuritkomplekser i berøringsfølsomme områder af huden, de findes i det basale lag af epidermis i fingre, læber og kønsorganer. De findes også i behåret hud med lavere tæthed. Merkel celle neuritkomplekset består af en Merkel celle i tæt position til en forstørret nerveterminal fra en enkelt myeliniseret A? fiber [fig. 1 (C1)] (gennemgang i Halata og samarbejdspartnere) .29 På den epidermale side udviser Merkel-celle fingerlignende processer, der strækker sig mellem nærliggende keratinocytter [fig. 1 (C2)]. Merkel-celler er keratinocyt-afledte epidermale celler.30,31 Touch touch dome blev introduceret for at navngive den store koncentration af Merkel-cellekomplekser i den hårede hud på kattepoten. En berøringskuppel kunne have op til 150 Merkel-celler, der er innerveret af en enkelt A? -Fiber og hos mennesker foruden A? -Fibre, A? og C-fibre var også regelmæssigt til stede. 32-34
Stimulering af Merkel-celleneuritkomplekser resulterer i langsomt tilpasning af Type I (SA I) -reaktioner, der stammer fra punkterede modtagelige felter med skarpe grænser. Der er ingen spontan udledning. Disse komplekser reagerer på fordybningens dybde og har den højeste rumlige opløsning (0.5 mm) af de kutane mekanoreceptorer. De transmitterer et præcist rumligt billede af taktile stimuli og foreslås at være ansvarlige for form og teksturdiskrimination [fig. 2 (B1)]. Mus blottet for Merkel-celler kan ikke registrere strukturerede overflader med deres fødder, mens de gør det ved hjælp af deres whiskers.35
Uanset om Merkel-cellen, den sensoriske neuron eller begge steder er mekanotransduktion, er det stadig et spørgsmål om debat. Hos rotter afskaffer fototoksisk ødelæggelse af Merkel-celler SA I-respons.36 Hos mus med genetisk undertrykkede Merkel-celler forsvandt SA I-svaret, der blev registreret i ex vivo hud / nervepræparat, fuldstændigt, hvilket viser, at Merkel-celler er nødvendige for korrekt kodning af Merkel receptorresponser.37 Den mekaniske stimulering af isolerede Merkel-celler i kultur ved motordrevet tryk genererer imidlertid ikke mekanisk styrede strømme.38,39 Keratinocytter kan spille en vigtig rolle i den normale funktion af Merkel-cellenneuritkomplekset. Merkel-cellefingerlignende processer kan bevæge sig med huddeformation og epidermis cellebevægelse, og dette kan være det første trin i mekanisk transduktion. Det er klart, at de betingelser, der kræves for at studere mechano-følsomhed af Merkel-celler, endnu ikke er etableret.
Ruffini slutninger. Ruffini-ender er tynde cigareformede indkapslede sensoriske ender forbundet med A? nerveender. Ruffini-ender er små bindevævscylindre arrangeret langs dermale kollagenstrenge, der leveres af en til tre myeliniserede nervefibre med en diameter på 4 mm. Op til tre cylindre med forskellig orientering i dermis kan smelte sammen og danne en receptor [fig. 6 (C1)]. Strukturelt ligner Ruffini-enderne Golgi senerorganer. De udtrykkes bredt i dermis og er blevet identificeret som de langsomt tilpasende kutane mekanoreceptorer af type II (SA II). På baggrund af spontan nervøs aktivitet fremkaldes en langsomt tilpasning af regelmæssig afladning ved vinkelret lav kraft, der opretholdes mekanisk stimulering eller mere effektivt ved hududstrækning. SA II-respons stammer fra store modtagelige felter med obskure grænser. Ruffini-receptorer bidrager til opfattelsen af retningen af objektbevægelse gennem mønsteret af hudstrækning [fig. 3 (A2)].
I mus kan SA I- og SA II-reaktioner adskilles elektrofysiologisk i ex-vivo nervehud-præparation. 40 Nandasena og samarbejdspartnere rapporterede immunokalokalisering af aquaporin 1 (AQP1) i de periodontale Ruffini-slutninger af rotteformerne, hvilket tyder på, at AQP1 er involveret i opretholdelsen af den dental osmotiske balance, der er nødvendig for mekanotransduktion. 41 De periodontale Ruffini-endinger udtrykte også den formodede mekanosensitive ionkanal ASIC3.42
Meissner-legemer. Meissner-legemer er lokaliseret i den glatte hudens dermale papiller, hovedsageligt i håndflader og fodsåler, men også i læber, tunge, ansigt, brystvorter og kønsorganer. Anatomisk består de af en indkapslet nerveende, hvor kapslen er lavet af flade understøttende celler arrangeret som vandrette lameller indlejret i bindevæv. Der er en enkelt nervefiber A? afferenter forbundne pr. legemsvæske [Fig. 1 (C4)]. Enhver fysisk deformation af kroppen viser en volley af handlingspotentialer, der hurtigt ophører, dvs. de tilpasser sig hurtigt receptorer. Når stimulus fjernes, genvinder kroppsformen sin form og producerer samtidig en ny volley af handlingspotentialer. På grund af deres overfladiske placering i dermis reagerer disse kroppe selektivt på hudbevægelse, taktil detektion af glidning og vibrationer (20 Hz). De er følsomme over for dynamisk hud - for eksempel mellem huden og en genstand, der håndteres [fig. 40 (A2)].
Pacinian corpuscles. Pacininske legemer er de dybere mekanoreceptorer i huden og er den mest følsomme indkapslede kutane mekanoreceptor for hudbevægelse. Disse store ovale blodlegemer (1 mm i længden) lavet af koncentriske lameller af fibrøst bindevæv og fibroblaster foret med fladt modificerede Schwann-celler udtrykkes i den dybe dermis.43 I midten af kroppen, i et væskefyldt hulrum kaldet indre pære , afslutter en enkelt A? afferent umyeliniseret nerveende [fig. 1 (C5)]. De har et stort modtageligt felt på hudens overflade med et særligt følsomt center. Udviklingen og funktionen af flere hurtigt tilpassende mekanoreceptortyper forstyrres i c-Maf-mutante mus. Især Pacininske kroppe er alvorligt atrofiere.44
Pacininske blodlegemer viser meget hurtig tilpasning som reaktion på fordybningen af huden, den hurtigt tilpasende II (RA II) nerveafladning, der er i stand til at følge høj frekvens af vibrerende stimuli, og tillader opfattelse af fjerne begivenheder gennem transmitterede vibrationer.45 Pacinian corpuscle afferenter reagerer på vedvarende fordybning med forbigående aktivitet ved stimulusens begyndelse og forskydning. De kaldes også accelerationsdetektorer, fordi de kan registrere ændringer i stimulusens styrke, og hvis ændringshastigheden i stimulus ændres (som det sker i vibrationer), bliver deres svar proportionalt med denne ændring. Paciniske legemer mærker grove trykændringer og mest af alt vibrationer (150 300 Hz), som de kan registrere selv centimeter væk [fig. 2 (A3)].
Tonisk respons blev observeret i decapsulated Pacinian corpuscle.46 Derudover reagerer intakte Pacinian-korpusker med vedvarende aktivitet under konstante indrykkestimuli uden at ændre mekaniske tærskler eller responsfrekvens, når GABA-medieret signalering er blokeret mellem lamellatglia og en nerveendring. 47. de ikke-neuronale komponenter i Pacinian-korpuslet kan have dobbelt roller i filtrering af den mekaniske stimulus såvel som til modulering af sensory neurons responsegenskaber.
Rygmarv fremspring. Fremskrivninger af A? -LTMR'erne i rygmarven er opdelt i to grene. Den vigtigste centrale gren stiger op i rygmarven i de ipsilaterale dorsale søjler til cervikal niveau [fig. 1 (B3)]. Sekundære grene slutter i ryghornet i laminerne IV og interfererer f.eks. Med smerteoverførslen. Dette kan dæmpe smerter som en del af portkontrollen [Fig. 1 (B4)]. 48
Ved cervicale niveauer separerer hovedpartens axoner i to områder: Midterbanen omfatter gracile fasciklen, der formidler information fra den nedre halvdel af kroppen (ben og stammen), og den ydre kanal omfatter den cuneate fascicle, der transporterer information fra den øvre halvdel af kroppen (arme og kuffert) [Fig. 1 (B5)].
Primære taktile afferenter laver deres første synaps med anden ordens neuroner ved medulla, hvor fibre fra hver kanal synapser i en kerne med samme navn: gracile fasciculus axones synaps i gracile nucleus og cuneat axones synapse i cuneatkernen [Fig. 1 (B6)]. Neuroner, der modtager synapsen, tilvejebringer de sekundære afferenter og krydser midtlinjen straks for at danne en kanal på den kontralaterale side af hjernestammen den mediale lemniscus som stiger gennem hjernestammen til den næste relæstation i midthjernen, specifikt i thalamus [Fig. . 1 (B7)].
Molekylær specifikation af LTMR'er. Molekylære mekanismer, der styrer den tidlige diversificering af LTMR'er, er for nylig blevet delvist belyst. Bourane og samarbejdspartnere har vist, at de neuronale populationer, der udtrykker Ret tyrosinkinasereceptoren (Ret) og dens co-receptor GFR? 2 i E11 13 embryonale mus, DRG selektivt udtrykker transkriptionsfaktoren Mafa.49,50 Disse forfattere viser, at Mafa / Ret / GFR2-neuroner bestemt til at blive tre specifikke typer LTRM'er ved fødslen: SA1-neuroner, der innerverer Merkel-cellekomplekser, de hurtigt tilpasende neuroner, der innerverer Meissner-legemer og de hurtigt tilpasende afferenter (RA I), der danner lancetformede ender omkring hårsækkene. Ginty og samarbejdspartnere rapporterer også, at DRG-neuroner, der udtrykker early-Ret, hurtigt tilpasser sig mekanoreceptorer fra Meissner-blodlegemer, Pacinian-blodlegemer og lancetformede ender omkring hårsækkene.51 De innerverer diskrete målzoner inden for gracile og cuneate kerner og afslører et modalitetsspecifikt mønster af mekanosensor neuronale aksonale fremspring i hjernestammen.
Undersøgelse af humane hudmekanoreceptorer. Teknikken til mikroneurografi beskrevet af Hagbarth og Vallbo i 1968 er blevet anvendt til at undersøge afladningsadfærden for enkelt menneskelige mekaniske følsomme ender, der forsyner muskel, led og hud (se til gennemgang Macefield, 2005). 52,53 Størstedelen af mikronurografi på huden undersøgelser har karakteriseret fysiologien af taktile afferenter i håndens glat hud. Mikroelektrodeoptagelser fra median- og ulnarnervene hos mennesker har afsløret berøringsfølelse genereret af de fire klasser af LTMR'er: Meissner-afferenter er særligt følsomme over for lysstrøg over huden og reagerer på lokale forskydningskræfter og begyndende eller åbenlyse glider inden i det modtagelige felt. Paciniske afferenter er udsøgte følsomme over for hurtige mekaniske transienter. Tilbudere reagerer kraftigt på at blæse over det modtagelige felt. Et Pacinian-legeme placeret i et ciffer vil normalt svare på at banke på bordet, der understøtter armen. Merkel-afferenter har karakteristisk en høj dynamisk følsomhed over for indrykningsstimuli anvendt på et diskret område og reagerer ofte med en afladning under frigivelse. Selvom Ruffini-afferenterne reagerer på kræfter, der påføres normalt på huden, er et unikt træk ved SA II-afferenter deres evne til at reagere også på lateral hudstrækning. Endelig har hårenheder i underarmen store ovale eller uregelmæssige modtagelige felter sammensat af flere følsomme pletter, der svarede til individuelle hår (hver afferent forsyning ~ 20 hår).
Keratinocytternes mekaniske følsomhed
Enhver mekanisk stimulering på huden skal overføres gennem keratinocytter, der danner epidermis. Disse allestedsnærværende celler kan udføre signaleringsfunktioner ud over deres støttende eller beskyttende roller. For eksempel udskiller keratinocytter ATP, et vigtigt sensorisk signalmolekyle som reaktion på mekaniske og osmotiske stimuli. 54,55 Udløsningen af ATP inducerer intracellulær calciumforøgelse ved autokrin stimulering af purinergreceptorer. 55 Derudover er der tegn på, at hypotonicitet aktiverer Rho-kinasen signalveje og den efterfølgende F-actin-spændingsfiberdannelse, hvilket tyder på, at den mekaniske deformation af keratinocytterne mekanisk kan interferere med nabo-cellerne, såsom Merkel-celler, til uskadelig berøring og C-fiberfri ende for skadelig berøring [Fig. 1 (C6)]. 56,57
Skadelig berøring
Høj tærskel mekanoreceptorer (HTMR'er) er epidermale C- og A? gratis nerveender. De er ikke forbundet med specialiserede strukturer og observeres i begge hårhud [fig. 1 (A9)] og glat hud [fig. 1 (C7)]. Imidlertid skal betegnelsen fri nerveendende betragtes med forsigtighed, da nerveender altid er i tæt placering med keratinocyt- eller Langherans-celle eller melanocytter. Ultrastrukturel analyse af nerveender afslører tilstedeværelsen af groft endoplasmatisk retikulum, rigelig mitokondrier og tætkerne-vesikel. Tilstødende membraner af epidermale celler fortykkes og ligner postsynaptisk membran i nervevæv. Bemærk, at interaktionerne mellem nerveender og epidermale celler kan være tovejs, da epidermale celler kan frigive mediatorer som ATP, interleukin (IL6, IL10) og bradykinin, og omvendt kan peptidergiske nerveender frigive peptider såsom CGRP eller substans P, der virker på epidermale celler. HTMR'er omfatter mechano-nociceptorer, der kun ophidses af skadelige mekaniske stimuli og polymodale nociceptorer, der også reagerer på skadelig varme og eksogent kemikalie [Fig. 2 (B2)] 58
HTMR-afferente fibre slutter på projektionsneuroner i rygmarvs dorsale horn. A? -HTMR'er kommer i kontakt med andenordens neuroner overvejende i lamina I og V, hvorimod C-HTMR'er afsluttes i lamina II [Fig. 1 (B8)]. Anden ordens nociceptive neuroner rager ud til den kontrolsidede side af rygmarven og stiger op i det hvide stof og danner det anterolaterale system. Disse neuroner slutter hovedsageligt i thalamus [fig. 1 (B9 og B10)].
Mechano-strømme i Somatosensoriske Neuroner
Mekanismerne til langsom eller hurtig tilpasning af mekanoreceptorer er endnu ikke klarlagt. Det er uklart, i hvilket omfang mekanoreceptortilpasning tilvejebringes af den cellulære miljø i den sensoriske nerveafslutning, de mekaniske gatedkanalers egenskaber og egenskaberne af de aksonale spændingsgatede ionkanaler i sensoriske neuroner (Fig. 2). De seneste fremskridt i karakteriseringen af mekanisk gatedstrømme har imidlertid vist, at forskellige klasser af mekanosensitive kanaler findes i DRG-neuroner og kan forklare nogle aspekter af tilpasningen af mekanoreceptorer.
In vitro-registrering hos gnavere har vist, at somaen fra DRG-neuroner er iboende mekanosensitiv og udtrykker kationiske mechano-gatede strømme.59-64 Gadolinium og rutheniumrød blokerer fuldt ud mekanosensitive strømme, hvorimod eksternt calcium og magnesium ved fysiologiske koncentrationer samt amilorid og benzamil, forårsager delvis blokering. 60,62,63 FM1-43 fungerer som en varig blokering, og injektionen af FM1-43 i musens bagpote nedsætter smertefølsomheden i Randall Selitto-testen og øger den vurderede tærskel for tilbagetrækning af poten med von Frey-hår.65
Som reaktion på vedvarende mekanisk stimulering falder mekanosensitive strømme gennem lukning. Baseret på tidskonstanterne for det nuværende forfald er der adskilt fire forskellige typer af mekanosensitive strømme: hurtigt tilpasning af strømme (~ 3 6 ms), mellemliggende tilpasning af strømme (~ 15 30 ms), langsomt tilpasning af strømme (~ 200 300 ms) ) og ultra-langsomt tilpasningsstrømme (~ 1000 ms) .64 Alle disse strømme er til stede med variabel forekomst i rotte-DRG-neuroner, der innerverer bagbenets glat hud.64
Den mekaniske følsomhed af mekanosensitive strømme kan bestemmes ved at anvende en række inkrementelle mekaniske stimuli, hvilket giver mulighed for relativt detaljeret stimulusstrømsanalyse.66 Stimulus strømsforholdet er typisk sigmoidalt, og den maksimale amplitude af strømmen bestemmes af antallet af kanaler, der er åbne samtidigt.64,67 Interessant nok er det blevet rapporteret, at den hurtigt tilpasende mekanosensitive strøm viser lav mekanisk tærskel og halvaktiverende midtpunkt sammenlignet med den ultra-langsomt tilpasende mekanosensitive strøm.63,65
Sensoriske neuroner med ikke-nociceptive fænotyper udtrykker fortrinsvis hurtig tilpasning af mekanosensitive strømme med lavere mekanisk tærskel. 60,61,63,64,68 Omvendt rapporteres langsomt og ultra-langsomt tilpasning af mekanosensitive strømme i formodede ikke-nociceptive celler. 64,68 Dette fremkaldte forslag om, at disse strømme kunne bidrage til forskellige mekaniske tærskler ses i LTMR'er og HTMR'er in vivo. Skønt disse in vitro-forsøg bør tages med forsigtighed, blev der også tilvejebragt støtte til tilstedeværelsen i DR-neuronernes neuroner af lav- og høj-tærskelmekanotransducere ved radial strækbaseret stimulation af dyrkede mus-sensoriske neuroner. 69 Dette paradigme afslørede to hovedpopulationer af strækningskänslige neuroner, en som reagerer på lav stimulus amplitude og en anden som selektivt reagerer på høj stimulus amplitude.
Disse resultater har vigtige, men spekulative, mekaniske implikationer: Den mekaniske tærskel af sensoriske neuroner kan have lidt at gøre med den mekaniske organisering af mekanoreceptoren, men kan ligge i egenskaberne af de mekanisk gatede ionkanaler.
Mekanismerne, der ligger til grund for desensibilisering af mekanosensitive kationstrømme i DRG-neuroner fra rotter, er for nylig blevet afsløret.64,67 Det skyldes to samtidige mekanismer, der påvirker kanalegenskaber: tilpasning og inaktivering. Tilpasning blev først rapporteret i auditive hårcelleundersøgelser. Det kan beskrives operationelt som en simpel oversættelse af transducerkanalens aktiveringskurve langs den mekaniske stimulusakse. 70-72 Tilpasning tillader sensoriske receptorer at opretholde deres følsomhed over for nye stimuli i nærværelse af en eksisterende stimulus. Imidlertid kan en væsentlig del af mekanosensitive strømme i DRG-neuroner ikke genaktiveres efter konditionering af mekanisk stimulering, hvilket indikerer inaktivering af nogle transducerkanaler.64,67 Derfor virker både inaktivering og tilpasning i tandem for at regulere mekanosensitive strømme. Disse to mekanismer er fælles for alle mekanosensitive strømme identificeret i rotte-DRG-neuroner, hvilket tyder på, at relaterede fysisk-kemiske elementer bestemmer kinetikken for disse kanaler.64
Afslutningsvis er det afgørende at bestemme egenskaberne af endogene mekanosensitive strømme in vitro i søgen efter at identificere transduktionsmekanismer på molekylært niveau. Variationen observeret i den mekaniske tærskel og tilpasningskinetikken for de forskellige mekanisk gatede strømme i DRG-neuroner antyder, at ionkanalernes indre egenskaber i det mindste delvist kan forklare mekanisk tærskel- og tilpasningskinetik hos de mekanoreceptorer, der er beskrevet i årtierne 1960 80 ved anvendelse af ex vivo-præparater.
Putative Mekanosensitive Proteiner
Mekanisk følsomme ionstrømme i somatosensoriske neuroner karakteriseres godt, derimod er der ikke meget kendt om identiteten af molekyler, som medierer mekanotransduktion hos pattedyr. Genetiske skærme i Drosophila og C. elegans har identificeret kandidatmekanotransduktionsmolekyler, herunder TRP- og degenerin / epithelial Na + -kanalen (Deg / ENaC) -familierne. 73 Nylige forsøg på at belyse molekylære grundlag for mekanotransduktion hos pattedyr har i høj grad fokuseret på homologer af disse kandidater . Derudover er mange af disse kandidater til stede i kutane mekanoreceptorer og somatosensoriske neuroner (Fig. 2).
Acid-Sensing Ion Channels
ASIC'er tilhører en proton-gated undergruppe af degenerin epithelial Na + kanalfamilien.74 Tre medlemmer af ASIC-familien (ASIC1, ASIC2 og ASIC3) udtrykkes i mekanoreceptorer og nociceptorer. ASIC-kanalers rolle er blevet undersøgt i adfærdsmæssige studier ved anvendelse af mus med målrettet sletning af ASIC-kanalgener. Sletning af ASIC1 ændrer ikke funktionen af kutane mekaniske receptorer, men øger afferenters mekaniske følsomhed, der innerverer tarmen.75 ASIC2 knockout-mus udviser en nedsat følsomhed for hurtigt tilpasning af kutane LTMR'er.76 Imidlertid rapporterede efterfølgende studier manglende virkninger af at slå ASIC2 ud på både visceral mechano-nociception og kutan mekanosensation.77 ASIC3-forstyrrelse mindsker mechano-følsomhed hos viscerale afferenter og reducerer responsen af kutane HTMR'er på skadelige stimuli.76
Den Transient Receptor Channel
TRP superfamilien er opdelt i seks subfamilier i pattedyr. 78 Næsten alle TRP-underfamilier har medlemmer knyttet til mekanosensation i en række cellesystemer. 79 I pattedyrs sensoriske neuroner er TRP-kanaler imidlertid bedst kendt for at detektere termisk information og formidlende neurogen inflammation, og kun to TRP-kanaler, TRPV4 og TRPA1, har været impliceret i kontaktfølsomhed. Forstyrrende TRPV4-ekspression hos mus har kun beskedne virkninger på akutte mekanosensoriske tærskler, men reducerer kraftigt følsomheden overfor skadelige mekaniske stimuli. 80,81 TRPV4 er en afgørende determinant i formgivning af nociceptive neurons reaktion på osmotisk stress og mekanisk hyperalgesi under betændelse. 82,83 TRPA1 synes at har en rolle i mekanisk hyperalgesi. TRPA1-mangelfulde mus udviser smerteoverfølsomhed. TRPA1 bidrager til transduktion af mekaniske, kolde og kemiske stimuli i nociceptor sensoriske neuroner, men det ser ud til, at det ikke er vigtigt for hårcelletransduktion. 84,85
Der er intet klart bevis på, at TRP-kanaler og ASIC-kanaler, der er udtrykt i pattedyr, er mekanisk gatede. Ingen af disse kanaler udtrykker heterologt rekapitulerer den elektriske signatur af mekanosensitive strømme observeret i deres native miljø. Dette udelukker ikke muligheden for, at ASIC'er og TRPs-kanaler er mekanotransducere, givet usikkerheden om, hvorvidt en mekanotransduktionskanal kan fungere uden for sin cellulære kontekst (se afsnit om SLP3).
Piezo Proteiner
Piezo protiens er for nylig blevet identificeret som lovende kandidater til mekanosenserende proteiner fra Coste og collaborators.86,87 Vertebrates har to Piezo-medlemmer, Piezo 1 og Piezo 2, der tidligere er kendt som FAM38A og FAM38B, som er godt konserveret gennem multi-cellulære eukaryoter. Piezo 2 er rigeligt i DRG'er, mens Piezo 1 næppe kan påvises. Piezo-inducerede mekanosensitive strømme forhindres hæmmet af gadolinium, rutheniumrød og GsMTx4 (et toksin fra tarantula Grammostola spatulata) .88 Ekspression af Piezo 1 eller Piezo 2 i heterologe systemer frembringer mekanosensitive strømme, kinetikken ved inaktivering af Piezo 2 strøm er hurtigere end Piezo 1. På samme måde som endogene mekanosensitive strømme har Piezo-afhængige strømme reverseringspotentialer omkring 0 mV og er kation-ingen selektive, med Na +, K +, Ca2 + og Mg2 +, der gennemsyrer den underliggende kanal. På samme måde reguleres piezoafhængige strømme af membranpotentiale med en markant bremsning af den nuværende kinetik ved depolariserede potentialer. 86
Piezo proteiner er utvivlsomt mekanosenserende proteiner og deler mange egenskaber ved hurtigt at tilpasse mekanosensitive strømme i sensoriske neuroner. Behandling af dyrkede DRG-neuroner med Piezo 2-kort interfererende RNA reducerede andelen af neuroner med hurtig tilpasning af strøm og nedsat procentdelen af mekanosensitive neuroner. 86 Transmembrane-domæner er placeret i hele piezo-proteinerne, men der er ingen indlysende poreholdige motiver eller ionkanal signaturer blevet identificeret. Mus Piezo 1-protein renset og rekonstitueret i asymmetriske lipid-bilayere og liposomer danner ionkanaler, der er følsomme for rutheniumrød. 87 Et vigtigt trin i validerende mekanotransduktion gennem Piezo-kanaler er at anvende in vivo-fremgangsmåder til at bestemme den funktionelle betydning i berøringssignalering. Oplysninger blev givet i Drosophila, hvor sletning af det enkelte Piezo-medlem reducerede mekanisk reaktion på skadelige stimuli uden at påvirke normal berøring. 89 Selvom deres struktur fortsat skal bestemmes, er denne nye familie af mekanosensitive proteiner et lovende emne til fremtidig forskning ud over grænsen af touch sensation. For eksempel viser en nylig undersøgelse af patienter med anæmi (arvelig xerocytose) Piezo 1's rolle ved opretholdelse af erytrocytvolumen homeostase.90
Transmembrane Channel-Like (TMC)
En nylig undersøgelse viser, at to proteiner, TMC1 og TMC2 er nødvendige for hårcellemekanotransduktion. 91 Arvelig døvhed på grund af TMC1-genmutation blev rapporteret hos mennesker og mus. 92,93 Tilstedeværelsen af disse kanaler var endnu ikke vist i det somatosensoriske system, men det ser ud til at være en god ledelse til at undersøge.
Stomatin-lignende Protein 3 (SLP3)
Ud over transduktionskanalerne har nogle tilbehørsproteiner forbundet med kanalen vist sig at spille en rolle i berøringsfølsomhed. SLP3 udtrykkes i mammale DRG neuroner. Undersøgelser, der anvendte mutante mus, der manglede SLP3, havde vist ændring i mekanosensation og mekanosensorrige strømme. 94,95 SLP3 præcise funktion forbliver ukendt. Det kan være en linker mellem den mekanosensitive kanal og de underliggende mikrotubuli, som foreslået for sin C. elegans homolog MEC2.96 Nyligt GR. Lewin lab har antydet, at en tether syntetiseres af DRG sensoriske neuroner og forbinder mekanosensitiv ionkanal med den ekstracellulære matrix. 97 Forstyrrende forbindelsen afskaffer den RA-mekanosensitive strøm, hvilket tyder på, at nogle ionkanaler kun er mekanisk følsomme, når de er tæmmet. RA-mekanosensitive strømme hæmmes også af laminin-332, et matrixprotein produceret af keratinocytter, der forstærker hypotesen af en modulering af den mekanosensitive strøm af ekstracellulære proteiner. 98
K + Kanalsubfamilie
Parallelt med kationiske depolariserende mekanosensitive strømme undersøges tilstedeværelsen af repolariserende mekanosensitive K + strømme. K + -kanaler i mekanosensitive celler kan træde i den aktuelle balance og bidrage til at definere den mekaniske tærskel og tidskurset for tilpasning af mekanoreceptorer.
KCNK medlemmer tilhører de to-pore domæne K + -kanal (K2P) family.99,100 The K2P vise en bemærkelsesværdig række regulering af cellulære, fysiske og farmakologiske midler, herunder pH-ændringer, varme, stræk og membran deformation. Disse K2P er aktive ved hvilemembranpotentiale. Adskillige KCNK underenheder udtrykkes i somatosensoriske neurones.101 KCNK2 (TREK-1), KCNK4 (TRAAK) og TREK-2 kanaler er blandt de få kanaler, som en direkte mekanisk gating ved membranfiltrering strækning har været shown.102,103
Mus med et forstyrret KCNK2-gen viste en forbedret følsomhed over for varme og milde mekaniske stimuli, men en normal tilbagetrækningstærskel for skadeligt mekanisk tryk på bagpoten ved anvendelse af Randall Selitto-testen.104 KCNK2-mangelfulde mus viser også øget termisk og mekanisk hyperalgesi ved inflammatorisk betingelser. KCNK4-knockout-mus var overfølsomme over for mild mekanisk stimulering, og denne overfølsomhed blev øget ved yderligere inaktivering af KCNK2.105 Øget mekanosensitivitet af disse knockout-mus kunne betyde, at stretch normalt aktiverer både depolarisering og repolarisering af mekanosensitive strømme på en koordineret måde, ligesom ubalancen mellem depolarisering og repolarisering af spændingsstyrede strømme.
KCNK18 (TRESK) er en vigtig bidragyder til baggrunden K + konduktans, der regulerer hvilemembranpotentialet for somatosensoriske neuroner.106 Selvom det ikke vides, om KCNK18 er direkte følsom over for mekanisk stimulering, kan det spille en rolle i formidling af reaktioner på let berøring, såvel som smertefulde mekaniske stimuli. KCNK18 og i mindre grad KCNK3 foreslås at være det molekylære mål for hydroxy -? - sanshool, en forbindelse fundet i Schezuan peberkorn, der aktiverer berøringsreceptorer og inducerer en prikkende fornemmelse hos mennesker. 107,108
Den spændingsafhængige K + kanal KCNQ4 (Kv7.4) er afgørende for at indstille hastigheden og frekvenspræferensen af en subpopulation af hurtigt tilpassende mekanoreceptorer i både mus og mennesker. Mutation af KCNQ4 er i første omgang forbundet med en form for arvelig døvhed. Interessant nok lokaliserer en nylig undersøgelse KCNQ4 i perifere nerveender af kutan hurtigt tilpasning af hårfollikel og Meissner corpuscle. Følgelig fører tab af KCNQ4-funktion til en selektiv forstærkning af mekanoreceptorfølsomhed over for lavfrekvent vibration. Navnlig viser personer med forsinket høretab på grund af dominerende mutationer i KCNQ4-genet forbedret ydeevne ved detektering af små amplitude, lavfrekvente vibrationer. 109

Dr. Alex Jimenez's Insight
Touch anses for at være en af de mest komplekse sanser i menneskekroppen, især fordi der ikke er noget specifikt organ der har ansvaret for det. I stedet opstår følelsesfølsomheden gennem sensoriske receptorer, kendt som mekanoreceptorer, som findes over huden og reagerer på mekanisk tryk eller forvrængning. Der findes fire hovedtyper af mekanoreceptorer i den glabroøse eller hårløse hud hos pattedyr: lamellære legemer, taktile legemer, Merkel-nerveendinger og pærekroppe. Mekanoreceptorer fungerer for at muliggøre detektion af berøring for at overvåge muskler, knogler og ledders position, kendt som proprioception, og endda at opdage lyde og bevægelse af kroppen. Forståelse af mekanismerne for struktur og funktion af disse mekanoreceptorer er et grundlæggende element i udnyttelsen af behandlinger og terapier til smertebehandling.
Konklusion
Touch er en kompleks forstand, fordi den repræsenterer forskellige taktile kvaliteter, nemlig vibrationer, form, tekstur, fornøjelse og smerte, med forskellige diskriminerende forestillinger. Indtil videre var korrespondancen mellem et berøringsorgan og den psykofysiske sans korrelativ, og klassespecifikke molekylmarkører er bare ved at komme frem. Udviklingen af gnaverprøver, der matcher mangfoldigheden af berøringsadfærd, er nu påkrævet for at lette fremtidig genomforskning. Brugen af mus, der mangler specifikke delmængder af sensoriske afferente typer, vil i høj grad lette identifikation af mekanoreceptorer og sensoriske afferente fibre forbundet med en bestemt berøringsmodalitet. Interessant nok åbner et nyt papir det vigtige spørgsmål om det genetiske grundlag for mekanosensoriske træk hos mennesker og foreslår, at enkeltgenmutation kan påvirke berøringsfølsomheden negativt. 110 Dette understreger, at patofysiologien af det menneskelige berøringsunderskud i vid udstrækning er ukendt og ville helt sikkert fremskridt ved at identificere nøjagtigt delmængden af sensoriske neuroner forbundet med en berøringsmodalitet eller et trykunderskud.
Til gengæld er der gjort fremskridt med at definere de biokysiske egenskaber ved de mechano-gated strømme. 64 Udviklingen af nye teknikker i de senere år, som muliggør overvågning af membranspændingsændringer, mens man registrerer mechano-gated strøm, har vist sig at være en værdifuld eksperimentel metode til at beskrive mekanosensitive strømme med hurtig, mellemliggende og langsom tilpasning (gennemgået i Delmas og samarbejdspartnere) .66,111 Fremtiden vil være at bestemme de nuværende egenskabers rolle i mekanismerne til tilpasning af funktionelt forskellige mekanoreceptorer og bidraget fra mekanosensitive K + strømme til excitabiliteten af LTMR'er og HTMR'er.
Den molekylære karakter af mechano-gated strømme i pattedyr er også et fremtidigt lovende forskningsemne. Fremtidig forskning vil fremskridt i to perspektiver, først for at bestemme rollen som tilbehørsmolekyle, der tinder kanaler til cytoskelettet og ville være forpligtet til at overføre eller regulere mekanosensitivitet af ionkanaler af lignende af TRP- og ASIC / EnaC-familier. For det andet at undersøge det store og lovende område af Piezo-kanalernes bidrag ved at besvare nøglespørgsmål i forhold til permeations- og gatingmekanismerne, delmængden af sensoriske neuroner og touchmodaliteter involverer Piezo og Piezo's rolle i ikke-neuronale celler associeret med mechanosensation.
Berøringssansen i sammenligning med synet, smagen, lyden og lugten, som bruger bestemte organer til at behandle disse fornemmelser, kan forekomme overalt i kroppen gennem små receptorer kendt som mekanoreceptorer. Forskellige typer mekanoreceptorer kan findes i forskellige lag af huden, hvor de kan opdage en bred vifte af mekanisk stimulering. Artiklen ovenfor beskriver specifikke højdepunkter, der demonstrerer fremskridt med strukturelle og funktionelle mekanismer hos mekanoreceptorer forbundet med berøringssansen. Information der henvises til fra National Center for Biotechnology Information (NCBI). Omfanget af vores information er begrænset til kiropraktik såvel som til rygmarvsskader og tilstande. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez

Yderligere emner: Rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og savnede dage på arbejdspladsen over hele verden. Faktisk er rygsmerter blevet tilskrevet som den næst mest almindelige årsag til doktorkontorbesøg, der kun overstiger luftvejsinfektioner. Ca. 80 procent af befolkningen vil opleve en form for rygsmerter mindst én gang i hele deres liv. Ryggraden er en kompleks struktur bestående af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette skader og / eller forværrede forhold, som f.eks herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.

EKSTRA VIGTIG TEMNE: Håndtering af lænderygsmerter
FLERE EMNER: EKSTRA EKSTRA:�Kroniske smerter og behandlinger
Blank
Referencer
Luk harmonika
Oplysningerne heri om "Strukturelle og funktionelle mekanismer af mekanoreceptorer" er ikke beregnet til at erstatte et en-til-en-forhold med en kvalificeret sundhedsperson eller autoriseret læge og er ikke medicinsk rådgivning. Vi opfordrer dig til at træffe sundhedsbeslutninger baseret på din forskning og partnerskab med en kvalificeret sundhedsperson.
Bloginformation og diskussioner om omfang
Vores informationsomfang er begrænset til kiropraktik, muskuloskeletal, fysisk medicin, wellness, bidragende ætiologisk viscerosomatiske forstyrrelser inden for kliniske præsentationer, tilhørende somatovisceral refleks klinisk dynamik, subluksationskomplekser, følsomme helbredsproblemer og/eller funktionel medicin artikler, emner og diskussioner.
Vi giver og præsenterer klinisk samarbejde med specialister fra forskellige discipliner. Hver specialist er styret af deres faglige omfang af praksis og deres licensjurisdiktion. Vi bruger funktionelle sundheds- og velværeprotokoller til at behandle og understøtte pleje af skader eller lidelser i bevægeapparatet.
Vores videoer, indlæg, emner, emner og indsigt dækker kliniske forhold, problemstillinger og emner, der relaterer til og direkte eller indirekte understøtter vores kliniske anvendelsesområde.*
Vores kontor har med rimelighed forsøgt at give støttende citater og har identificeret den eller de relevante forskningsundersøgelser, der understøtter vores indlæg. Vi leverer kopier af understøttende forskningsundersøgelser tilgængelige for tilsynsråd og offentligheden efter anmodning.
Vi forstår, at vi dækker forhold, der kræver yderligere forklaring på, hvordan det kan hjælpe med en bestemt plejeplan eller behandlingsprotokol. derfor er du velkommen til at spørge for yderligere at diskutere emnet ovenfor Dr. Alex Jimenez, DC, eller kontakte os på 915-850-0900.
Vi er her for at hjælpe dig og din familie.
Blessings
Dr. Alex Jimenez A.D. MSACP, RN*, CCST, Ifmcp*, CIFM*, ATN*
Email: coach@elpasofunctionalmedicine.com
Licenseret som Doctor of Chiropractic (DC) i Texas & New Mexico*
Texas DC-licensnummer TX5807, New Mexico DC Licensnr. NM-DC2182
Licenseret som registreret sygeplejerske (RN*) in Florida
Florida-licens RN-licens # RN9617241 (Kontrol nr. 3558029)
Kompakt status: Multi-State Licens: Bemyndiget til at praktisere i 40 stater*
Dr. Alex Jimenez DC, MSACP, RN*CIFM*, IFMCP*, ATN*, CCST
Mit digitale visitkort





