Rygklinik Oxidativ Stress Kiropraktik og Funktionel Medicin Team. Oxidativ stress er defineret som en forstyrrelse i balancen mellem produktionen af reaktivt ilt (frie radikaler) og antioxidantforsvar. Det er med andre ord en ubalance mellem produktionen af frie radikaler og kroppens evne til at modvirke eller afgifte de skadelige effekter gennem neutralisering af antioxidanter. Oxidativ stress fører til mange patofysiologiske tilstande i kroppen. Disse omfatter neurodegenerative sygdomme, dvs. Parkinsons sygdom, Alzheimers sygdom, genmutationer, kræftformer, kronisk træthedssyndrom, skrøbeligt X-syndrom, hjerte- og blodkarsygdomme, aterosklerose, hjertesvigt, hjerteanfald og inflammatoriske sygdomme. Oxidation sker under en række omstændigheder:
cellerne bruger glukose til at lave energi
immunsystemet bekæmper bakterier og skaber betændelse
kroppene afgifter forurenende stoffer, pesticider og cigaretrøg
Der foregår millioner af processer i vores kroppe på ethvert givet tidspunkt, som kan resultere i oxidation. Her er et par symptomer:
Træthed
Hukommelsestab og eller hjernetåge
Muskel- og eller ledsmerter
Rynker sammen med gråt hår
Nedsat syn
Hovedpine og følsomhed over for støj
Modtagelighed for infektioner
At vælge økologiske fødevarer og undgå toksiner i dit miljø gør en stor forskel. Dette, sammen med at reducere stress, kan være gavnligt til at reducere oxidation.
Vågner du træt, selv efter at have fået seks eller flere timers søvn?
Under en høj mængde stress?
Hvis du oplever nogen af disse situationer, kan det skyldes, at dine melatonin- og kortisolniveauer påvirker din krop og døgnrytme.
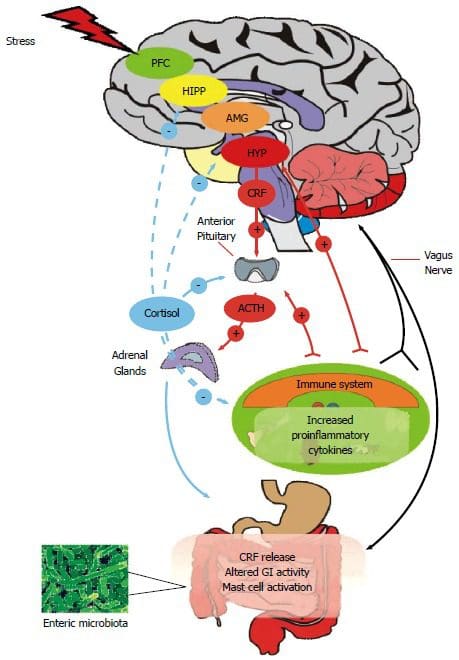
Over hele verden har millioner af mennesker problemer med at sove. I USA er der nogenlunde ca 50-70 millioner mennesker som har en dårlig søvnkvalitet. Når en person har sovet mindre end otte timer, bliver de trætte, og mange problemer kan komme til dem, især hvis deres liv er hektisk. Med en hektisk livsstil og dårlig søvn kan det medføre, at kroppen har lav energi til at få løst enhver opgave, kortisol stresshormonet vil blive hævet, og sygdomme som forhøjet blodtryk og diabetes kan give problemer, der kan være kroniske, hvis det ikke er tilfældet. behandlet.
I funktionel endokrinologi er melatonin og kortisol hormoner, som kroppen producerer naturligt. Kortisolhormonet eller stresshormonet hjælper kroppen med at være i en tilstand af "fight or flight", hvilket kan være en god ting for alle, der laver et projekt eller skal til jobsamtale. Selvom cortisolhormonniveauer er høje, kan det føre til, at kroppen får komplikationer som betændelse, kronisk oxidativ stress og højt blodtryk.
Melatonin døgnrytme
Med melatoninhormonet fortæller dette hormon kroppen, hvornår det er tid til at sove. Nogle gange har folk dog svært ved at sove, og at tage melatonintilskud kan faktisk slappe af i kroppen og dermed få personen til at falde i søvn. Da pinealkirtlen producerer melatonin fra hjernen, kan det også findes i øjnene, knoglemarven og tarmen for at slappe af i kroppen og få personen til at falde i søvn naturligt. Nogle studier viser at pinealkirtlens døgnrytme, der producerer melatonin. Ved at gøre dette viser forskningen, at administration af melatonin kan:
Én: fremkalde søvn hos personer, der har problemer med at falde i søvn.
to: hæmmer kroppen i at vågne naturligt fra den cirkadiske pacemaker.
Tre: Skift de cirkadiske biologiske ure for at øge søvnindtaget, når en person forsøger at sove på et tidligere tidspunkt for at få de fulde otte-timers fordele ved søvn.
Når en person arbejder på et 9 til 5 job, rejser de sig med deres kroppe og slapper af efter en hård dag på arbejde. Det fandt undersøgelser ud af at melatonin- og kortisolhormonerne er med til at regulere 24-timers mønsteret af kroppens funktion og reaktioner gevaldigt. Med kroppens hormonproduktionscyklus kan den forstyrres, hvis personen holder sig vågen sent om natten eller sover om dagen. Når dette sker, kan personen få forstyrrende lidelser som humørsvingninger, svimmelhed, være irritabel og deprimeret og have stofskifteforstyrrelser. Ikke kun det, men kroppens immunsystem og dets endokrine system kan også blive beskadiget, hvilket får kroppen til at være vært for infektioner og sygdomme.
Der har været flere undersøgelser af døgnrytmerne i kroppen, som viser undersøgelserne hvordan mennesker, der arbejder i nattevagt, er blevet forbundet med et stort antal ugunstige helbredsproblemer, der angriber det kardiovaskulære og mave-tarmsystem samt forstyrrer stofskiftet. Enhver, der har arbejdet nattevagt, er nødt til at ændre deres søvnplan og tilpasse sig den hurtige omlægning i deres søvn-/vågentidsplan for at gå på arbejde og udføre deres arbejde. Da alle arbejder på en vagtplan, kan det være stressende og kan påvirke en arbejders kropspræstation samt påvirke melatonin- og kortisolsekretionen.
Måder at støtte kortisol og melatonin på
Overraskende nok er der dog måder at sænke kortisolniveauerne på og sikre, at melatoninniveauet fungerer korrekt, for at kroppen kan fungere. For at kortisolniveauet skal sænkes, bør en person lave meditative øvelser, finde en fornøjelig hobby og, vigtigst af alt, prøve dybe vejrtrækningsøvelser for at slappe af kroppen fra uønsket stress. Med dybe vejrtrækningsøvelser kan det hjælpe kroppen med at frigøre enhver spænding, som en person holder, og musklerne i kroppen begyndte at slappe af, og blodet begynder at strømme. Med melatoninniveauerne arbejder de sammen med kroppens døgnrytme og sørger for, at kroppen ved, hvornår det er tid til at vågne, sove og spise. Melatoninhormonet kan også hjælpe med at regulere kroppens temperatur, blodtryk, og hormonniveauer for at sikre, at det fungerer korrekt. Når der er høje niveauer af disse systemer, kan det få kroppen til at udvikle kroniske sygdomme og skade kroppen i processen.
Forskning viser at melatoninhormoner kan binde sig til neurologiske receptorer i kroppen og dermed fremme afslapning. Da melatonin binder sig til neurologiske receptorer, kan det også reducere nerveaktivitet og dopaminniveauer for at gøre øjnene tunge og dermed få personen til at falde i søvn.
Konklusion
Med kroppen er i stand til naturligt at producere melatonin og cortisol niveauer for at sikre, at kroppen ikke bliver alt for stresset gennem hele dagen. Da melatonin er sammen med kroppens døgnrytme, ved kroppen, hvornår den skal holde sig oppe og falde i søvn. Da alle har et hektisk skema, er det vigtigt at tage sig tid og slappe af og få en sund søvnplan, så kroppen kan være sund og velfungerende. Nogle produkter er her for at sikre, at det endokrine system fungerer korrekt og understøtter binyrerne og sukkerstofskiftet.
Omfanget af vores information er begrænset til kiropraktik, muskuloskeletale og nervesundhedsmæssige problemer eller artikler, emner og diskussioner om funktionel medicin. Vi bruger funktionelle sundhedsprotokoller til at behandle skader eller lidelser i bevægeapparatet. Vores kontor har gjort et rimeligt forsøg på at give støttende citater og har identificeret den relevante forskningsundersøgelse eller undersøgelser, der understøtter vores stillinger. Vi stiller også kopier af understøttende forskningsundersøgelser til rådighed for bestyrelsen og eller offentligheden efter anmodning. For yderligere at diskutere emnet ovenfor, er du velkommen til at spørge Dr. Alex Jimenez eller kontakte os på 915-850-0900.
Referencer:
Cajochen, C, et al. �Melatonins rolle i reguleringen af menneskelige døgnrytmer og søvn.� Journal of Neuroendocrinology, US National Library of Medicine, april 2003, www.ncbi.nlm.nih.gov/pubmed/12622846.
James, Francine O, et al. � Cirkadiske rytmer af melatonin, kortisol og urgenekspression under simuleret natholdsarbejde.� Sleep, Associated Professional Sleep Societies, LLC, nov. 2007, www.ncbi.nlm.nih.gov/pmc/articles/PMC2082093/.
Monteleone, P, et al. �Tidsmæssigt forhold mellem melatonin og kortisol reaktioner på fysisk stress om natten hos mennesker.� Psychoneuroendocrinology, US National Library of Medicine, 1992, www.ncbi.nlm.nih.gov/pubmed/1609019.
Raman, Ryan. �Hvordan Melatonin kan hjælpe dig med at sove og få det bedre.� Healthline, Healthline Media, 3. september 2017, www.healthline.com/nutrition/melatonin-and-sleep.
Zamanian, Zahra, et al. �Oversigt over ændringer i cortisol og melatonin døgnrytme i sikkerhedsvagterne på Shiraz University of Medical Sciences.� International Journal of Preventive Medicine, Medknow Publications & Media Pvt Ltd, juli 2013, www.ncbi.nlm.nih.gov/pmc/articles/PMC3775223/.
Ved at informere enkeltpersoner om, hvordan National University of Health Sciences leverer viden til kommende generationer, der ønsker at gøre en forskel i verden. Universitetet tilbyder en bred vifte af medicinske erhverv til funktionel og integrerende medicin.
Har du nogensinde spekuleret på, hvorfor du føler dig træg efter en lang dag? Eller får du ondt i maven, når du har spist noget dårligt eller forkælet din yndlingsmad? Kan det være, at din tarm viser tegn på stress og ubehag på grund af visse vaner, som du kan støde på og ikke engang vidste om det?
I vores tidligere artikel talte vi om de seks typer mad at vores tarm skal være sund. Siden vores tarm indeholder billioner af mikrobiomer, både gode og dårlige, disse mikrobiomer spiller en vigtig rolle i vores generelle sundhed. Et sundt mikrobiom forbedrer vores gut sundhed, hjertesundhed, hjerne sundhed, styrer vores vægt , regulerer vores blodsukker. Med de gode bakterier i vores tarm, gavner bakterierne os med et godt fordøjelsessystem og ødelægger de skadelige bakterier. Men visse livsstile og kostvalg kan faktisk øge de dårlige bakterier og sænke de gode bakterier og det generelle helbred.
Her er fem overraskende livsstilsvalg, der skader din tarm:
Ikke spise en bred vifte af fødevarer
Vores tarm spiller en vigtig rolle i vores generelle sundhed. Når vi spiser gode hele fødevarer, er vores tarm gladere; vi har mere energi til at fuldføre enhver opgave, der bliver kastet på os, og vi får næringsstoffer til vores tarmflora. Men i løbet af de sidste par årtier har vi lænet os mere op i forarbejdede fødevarer på grund af det økonomiske pres fra øget fødevareproduktion. FOA udtalte, at 75 procent af verdens mad kun stammer fra 12 planter og fem dyrearter, og det er meget dårligt for vores tarmflora.
Her på Injury Medical & Chiropractic Clinic informerer vi vores patienter om vigtigheden af at spise nærende, hele fødevarer for ikke kun at fremme en sund tarm, men et sundt sind. Når kroppen bliver introduceret til en bred vifte af hele fødevarer (med et højt fiberindhold), begynder vores tarm at reparere skaderne på forarbejdet mad, som vi måske har indtaget internt.
Men når du ser bort fra præbiotika til din kost, er du det skade din fordøjelsessundhed. Uden præbiotika bremser vores fordøjelsessystem udviklingen og diversiteten for vores tarmflora. Så for at have en sundt mikrobiom udvikling, skal du inkorporere fødevarer fyldt med både fordøjelige og ufordøjelige fibre i din kost. Nogle fødevarer inkluderet i denne kategori er havre, nødder, løg, hvidløg, porrer, asparges, bananer, pærer, kikærter og bønner.
Det er måske en udfordring at holde sig til en kost med højt fiberindhold, men der er mulighed for at tage præbiotiske kosttilskud. Hvis du har et fødevareallergen eller fødevarefølsomhed over for fødevarer med højt fiberindhold, skal du tage præbiotiske kosttilskud kan faktisk hjælpe med at dyrke Bifidobacterium og Faecalibacterium i din tarm og være gavnlig for dit helbred uden ubehag.
For stort alkoholforbrug
Enhver voksen nyder alkohol en gang imellem. Ja, det er en af de drikkevarer, der hjælper dig med at slappe lidt af efter en lang dag, men for meget af det kan føre til alkoholmisbrug og afhængighed. Så vidste du, at det er dårligt for at indtage så meget alkohol dit hjerte, lever og hjerne; dermed skade din tarmsundhed og give dig dysbiose?
En undersøgelse anførte, at alkoholikere med dysbiose havde en lavere median forekomst af Bacteroidetes og en høj forekomst af Proteobakterier. Dem, der ikke var alkoholikere, blev ikke påvirket af undersøgelsen.
Imidlertid; der er nogle gode nyheder om at begrænse dig til alkoholisme, og at det kan være gavnligt for dine tarmbakterier. Hvis du moderat indtaget rødvin ansvarligt, polyfenoler i vinen kan være med til at gavne din tarmflora. Så nyd et glas vin en gang imellem som en lille forkælelse, der ikke bør tages for givet.
Utilstrækkelig søvn
I en af de tidligere artikler talte vi om, hvordan man opnår en god nats søvn gennem urter. Når vi får lidt til ingen søvn gennem vores hektiske liv, påvirker det os gennem forskellige helbredsproblemer, bl.a hjertesygdom , fedme. I en 2016 undersøgelse, opdagede forskere effekten af kortvarig søvnmangel på tarmmikrobiotaen efter to dage.
Når vores krop ikke får de anbefalede 8 timers søvn, tager vores tarm en kæmpe vejafgift, da vi føler os trætte og udmattede. Så for at sikre, at vores tarmmikrobiome vil blive taget hånd om, anbefaler vi at slukke for dine elektroniske enheder mindst 30 minutter før du gør dig klar til at slå dig ned for natten. Sluk alt lyset, og drik ikke væske mindst to timer før sengetid, luk øjnene og tag en dyb indånding i en meditativ tilstand, og slap af, mens du glider ind i slumrebyen.
Utilstrækkelig motion
Gennem vores hurtige livsstil og stressende job er det svært at finde tid til at træne. Men når vi rent faktisk finder tid til at træne, har vores sind det ikke kun godt; men vores krop og tarm har det også godt. Der dukker dog altid ting op, når vi er i en træningsrutine, og vi må springe træningen helt over. Det sker for os alle, og det er svært at fortsætte, hvor vi slap, da vi prøvede at træne.
Når vi ikke træner mindst et par gange om ugen, tager vores kroppe en enorm vejafgift på os, da vi tog på i vægt, vores stress er alt for højt, og vi har en højere chance at få en kronisk sygdom. Når dette sker, er vores tarmflora en stor ulempe. Her på klinikken stræber vi efter at informere vores patienter om vigtigheden af at træne, og at det ikke kun ændrer deres liv, men også ændrer deres humør totalt.
Du skal dog ikke bare gå ind i en hård træningsrutine, hvor du kommer til at skade dig selv. Start med en lavintensiv træning, og byg den derefter op, mens du går, fordi din tarmflora vil takke dig for det.
Som sidste ord ønsker vi her hos Injury Medical at holde dig orienteret om ernæring og måder at hjælpe dig med at forbedre dine lidelser med disse 5 overraskelser. Men også for at uddanne dig om, hvad der kan skade din tarm. Med disse overraskelser og små ændringer i dit daglige liv, vil din tarm takke dig for det lange seje.
NCBI Resources
Ifølge beviser fra et forskningsstudie fra 2016, tarmens immunsystem er grundlæggende for at forebygge en række sygdomme, og det kan ofte bidrage til stofskiftesygdomme. Det kan dog også hjælpe med at give et behandlingsmål, når man observerer systemisk inflammation i insulinresistens. Desuden er modificeret tarmimmunitet blevet forbundet med ændringer i tarmmikrobiotaen, tarmbarrierefunktionen, immunceller, der bor i tarmen, og resistens over for antigener, der trænger ind i mave-tarm- eller GI-systemet. Selvom dette tidligere har været antaget at øge faren for spiserørslidelser, herunder patogene infektioner og kronisk inflammation, som i sidste ende kan føre til kroniske helbredsproblemer.
Ketonlegemer skabes af leveren og bruges som energikilde, når glukose ikke er let tilgængelig i menneskekroppen. De to vigtigste ketonlegemer er acetoacetat (AcAc) og 3-beta-hydroxybutyrat (3HB), mens acetone er det tredje og mindst udbredte ketonlegeme. Ketoner er altid til stede i blodet, og deres niveauer stiger under faste og langvarig træningketogenese er den biokemiske proces, hvormed organismer producerer ketonlegemer gennem nedbrydning af fedtsyrer og ketogene aminosyrer.
Ketonlegemer skabes hovedsageligt i mitokondrier af leverceller. Ketogenese opstår, når der er lave glukoseindhold i blodet, især efter at andre cellulære kulhydratforretninger, såsom glykogen, er blevet opbrugt. Denne mekanisme kan også forekomme, når der ikke er tilstrækkelige mængder insulin. Fremstillingen af ketonlegemer er i sidste ende indledt for at stille tilgængelig energi til rådighed, som er lagret i den menneskelige krop som fedtsyrer. Ketogenese forekommer i mitokondrier, hvor det er uafhængigt reguleret.
Abstrakt
Ketonlegemetabolisme er en central knudepunkt i fysiologisk homeostase. I denne gennemgang diskuteres, hvordan ketoner tjener diskrete finjusterende metaboliske roller, der optimerer orgel og organismer ydeevne i varierende næringsstoffer forbliver og beskytter mod betændelse og skade i flere organsystemer. Traditionelt betragtes som metaboliske substrater, der kun er inddraget i kulhydratrestriktioner, understreges de seneste observationer betydningen af ketonlegemer som vigtige metaboliske og signalmediatorer, når kulhydrater er rigelige. Som supplement til et repertoire af kendte terapeutiske muligheder for sygdomme i nervesystemet er der opstået potentielle roller for ketonlegemer i kræft, som har spændende beskyttelsesroller i hjerte og lever, der åbner terapeutiske muligheder i fedme-relateret og hjerte-kar-sygdomme. Kontroverser i ketonmetabolisme og signalering diskuteres for at forene klassisk dogma med nutidige observationer.
Introduktion
Ketonlegemer er en vital alternativ metabolisk brændstofkilde for alle livets domæner, eukarya, bakterier og archaea (Aneja et al., 2002; Cahill GF Jr, 2006; Krishnakumar et al., 2008). Ketonlegememetabolisme hos mennesker er blevet udnyttet til at give næring til hjernen under episodiske perioder med mangel på næringsstoffer. Ketonlegemer er sammenvævet med vigtige pattedyrs metaboliske veje, såsom β-oxidation (FAO), tricarboxylsyrecyklussen (TCA), gluconeogenese, de novo lipogenese (DNL) og biosyntese af steroler. Hos pattedyr produceres ketonstoffer overvejende i leveren fra FAO-afledt acetyl-CoA, og de transporteres til ekstrahepatisk væv til terminal oxidation. Denne fysiologi giver et alternativt brændstof, der forstærkes af relativt korte perioder med faste, hvilket øger fedtsyretilgængeligheden og mindsker kulhydrattilgængeligheden (Cahill GF Jr, 2006; McGarry og Foster, 1980; Robinson og Williamson, 1980). Ketonlegemeoxidation bliver en væsentlig bidragyder til pattedyrs overordnede metabolisme i ekstrahepatisk væv i et utal af fysiologiske tilstande, herunder faste, sult, neonatalperioden, post-motion, graviditet og overholdelse af lavkulhydratdiæter. Cirkulerende samlede ketonlegemekoncentrationer hos raske voksne mennesker udviser normalt døgnsvingninger mellem ca. 100�250�M, stiger til ~1 mM efter langvarig træning eller 24 timers faste og kan akkumuleres til så højt som 20 mM i patologiske tilstande som diabetisk ketoacidose ( Cahill GF Jr, 2006; Johnson et al., 1969b; Koeslag et al., 1980; Robinson og Williamson, 1980; Wildenhoff et al., 1974). Den menneskelige lever producerer op til 300 g ketonlegemer om dagen (Balasse og Fery, 1989), som bidrager med mellem 5-20 % af det samlede energiforbrug i tilstande med mad, faste og sult (Balasse et al., 1978; Cox et al. al., 2016).
Nylige undersøgelser fremhæver nu vigtige roller for ketonlegemer i pattedyrcellemetabolisme, homeostase og signalering under en lang række fysiologiske og patologiske tilstande. Udover at fungere som energibrændstoffer til ekstrahepatiske væv som hjerne-, hjerte- eller skeletmuskulatur, spiller ketonlegemer hovedrolle som signalmediatorer, drivkræfter for protein post-translationel modifikation (PTM) og modulatorer af inflammation og oxidativ stress. I denne anmeldelse giver vi både klassisk og moderne syn på de pleiotropiske roller af ketonlegemer og deres metabolisme.
Oversigt over Ketonlegemetabolisme
Hastigheden af hepatisk ketogenese styres af en orkestreret række fysiologiske og biokemiske transformationer af fedt. Primære regulatorer inkluderer lipolyse af fedtsyrer fra triacylglyceroler, transport til og på tværs af hepatocytplasmamembranen, transport ind i mitokondrier via carnitin palmitoyltransferase 1 (CPT1), a-oxidationsspiralen, TCA-cyklusaktivitet og mellemkoncentrationer, redoxpotentiale og de hormonelle regulatorer af disse processer, overvejende glucagon og insulin [gennemgået i (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al., 1983; Kahn et al., 2005; McGarry and Foster , 1980; Williamson et al., 1969)]. Klassisk betragtes ketogenese som en overløbsvej, hvor a-oxidationsafledt acetyl-CoA overstiger citratsyntaseaktivitet og / eller oxaloacetat-tilgængelighed for kondens til dannelse af citrat. Mellemprodukter med tre carbon udviser anti-ketogen aktivitet, formodentlig på grund af deres evne til at udvide oxaloacetatpuljen til acetyl-CoA-forbrug, men hepatisk acetyl-CoA-koncentration alene bestemmer ikke ketogen hastighed (Foster, 1967; Rawat og Menahan, 1975; Williamson et al., 1969). Reguleringen af ketogenese ved hormonelle, transkriptionelle og post-translationelle begivenheder understøtter sammen tanken om, at de molekylære mekanismer, der finjusterer ketogen hastighed, forbliver ufuldstændigt forstået (se regulering af HMGCS2 og SCOT / OXCT1).
Ketogenese forekommer primært i lever mitokondrie matrix ved hastigheder, der er proportionale med total fedtoxidation. Efter transport af acylkæder over mitokondrie-membranerne og a-oxidation katalyserer mitokondrie-isoformen af 3-hydroxymethylglutaryl-CoA-syntase (HMGCS2) den skæbne, der begår kondensation af acetoacetyl-CoA (AcAc-CoA) og acetyl-CoA for at generere HMG-CoA (Fig. 1A). HMG-CoA lyase (HMGCL) spalter HMG-CoA for at frigøre acetyl-CoA og acetoacetat (AcAc), og sidstnævnte reduceres til d - a - hydroxybutyrat (d-? OHB) ved phosphatidylcholin-afhængig mitokondrie d-a OHB dehydrogenase ( BDH1) i en NAD + / NADH-koblet næsten ligevægtsreaktion (Bock og Fleischer, 1975; LEHNINGER et al., 1960). BDH1-ligevægtskonstant favoriserer d-? OHB-produktion, men forholdet mellem AcAc / d-? OHB-ketonlegemer er direkte proportionalt med mitokondrie NAD + / NADH-forhold, og således modulerer BDH1 oxidoreduktaseaktivitet mitokondrie redox potentiale (Krebs et al., 1969; Williamson et al., 1967). AcAc kan også spontant decarboxylere til acetone (Pedersen, 1929), kilden til sød lugt hos mennesker, der lider af ketoacidose (dvs. samlede serumketonlegemer> ~ 7 mM; AcAc pKa 3.6,? OHB pKa 4.7). Mekanismerne, gennem hvilke ketonlegemer transporteres gennem den mitokondriale indre membran, er ikke kendt, men AcAc / d-a-OHB frigøres fra celler via monocarboxylattransportører (hos pattedyr, MCT 1 og 2, også kendt som opløsningsbærer 16A-familiemedlemmer 1 og 7) og transporteres i cirkulationen til ekstrahepatiske væv til terminal oxidation (Cotter et al., 2011; Halestrap og Wilson, 2012; Halestrap, 2012; Hugo et al., 2012). Koncentrationer af cirkulerende ketonlegemer er højere end i ekstrahepatiske væv (Harrison og Long, 1940), hvilket indikerer, at ketonlegemer transporteres ned i en koncentrationsgradient. Funktionstabsmutationer i MCT1 er forbundet med spontane anfald af ketoacidose, hvilket antyder en kritisk rolle i ketonlegemeimport.
� Med undtagelse af potentiel omledning af ketonstoffer til ikke-oxidative skæbner (se Ikke-oxidative metaboliske skæbner for ketonstoffer), mangler hepatocytter evnen til at metabolisere de ketonstoffer, de producerer. Ketonlegemer syntetiseret de novo af leveren er (i) kataboliseret i mitokondrier i ekstrahepatisk væv til acetyl-CoA, som er tilgængelig for TCA-cyklussen til terminal oxidation (fig. 1A), (ii) omdirigeret til lipogenese- eller sterolsyntesevejene ( Fig. 1B), eller (iii) udskilles i urinen. Som et alternativt energetisk brændstof oxideres ketonlegemer ivrigt i hjerte, skeletmuskulatur og hjerne (Balasse og Fery, 1989; Bentourkia et al., 2009; Owen et al., 1967; Reichard et al., 1974; Sultan, 1988 ). Ekstrahepatisk mitokondriel BDH1 katalyserer den første reaktion af OHB-oxidation og omdanner den til back-AcAc (LEHNINGER et al., 1960; Sandermann et al., 1986). En cytoplasmatisk d-OHB-dehydrogenase (BDH2) med kun 20 % sekvensidentitet med BDH1 har en høj Km for ketonstoffer og spiller også en rolle i jernhomeostase (Davuluri et al., 2016; Guo et al., 2006) . I ekstrahepatisk mitokondriel matrix aktiveres AcAc til AcAc-CoA gennem udveksling af en CoA-del fra succinyl-CoA i en reaktion katalyseret af en unik pattedyrs CoA-transferase, succinyl-CoA:3-oxoacid-CoA-transferase (SCOT, CoA-transferase; kodet af OXCT1), gennem en næsten ligevægtsreaktion. Den frie energi frigivet ved hydrolyse af AcAc-CoA er større end den af succinyl-CoA, hvilket fremmer AcAc-dannelse. Således opstår ketonlegemets oxidative flux på grund af massevirkning: en rigelig forsyning af AcAc og hurtigt forbrug af acetyl-CoA gennem citratsyntase fremmer AcAc-CoA (+ succinat) dannelse af SCOT. Især, i modsætning til glucose (hexokinase) og fedtsyrer (acyl-CoA-syntetaser), kræver aktivering af ketonstoffer (SCOT) til en oxiderbar form ikke investering af ATP. En reversibel AcAc-CoA thiolasereaktion [katalyseret af en af de fire mitokondrielle thiolaser kodet af enten ACAA2 (koder for et enzym kendt som T1 eller CT), ACAT1 (koder for T2), HADHA eller HADHB] giver to molekyler af acetyl-CoA, som indgår i TCA-cyklussen (Hersh og Jencks, 1967; Stern et al., 1956; Williamson et al., 1971). Under ketotiske tilstande (dvs. totale serumketoner > 500 �M) bliver ketonlegemer væsentlige bidragydere til energiforbruget og udnyttes hurtigt i væv, indtil optagelse eller mætning af oxidation finder sted (Balasse et al., 1978; Balasse og Fery, 1989 Edmond et al., 1987). En meget lille del af leverafledte ketonstoffer kan let måles i urinen, og udnyttelse og reabsorptionshastigheder i nyrerne er proportionale med cirkulerende koncentration (Goldstein, 1987; Robinson og Williamson, 1980). Under stærkt ketotiske tilstande (> 1 mM i plasma) tjener ketonuri som en semi-kvantitativ reporter af ketose, selvom de fleste kliniske assays af urinketonlegemer påviser AcAc, men ikke ?OHB (Klocker et al., 2013).
Ketogene substrat og deres indvirkning på hepatocyt metabolisme
Ketogene substrater indbefatter fedtsyrer og aminosyrer (fig. 1B). Katabolismen af aminosyrer, især leucin, genererer omkring 4% af ketonlegemer i postabsorberende tilstand (Thomas et al., 1982). Således er acetyl-CoA-substratpuljen til dannelse af ketonlegemer hovedsagelig afledt af fedtsyrer, fordi pyruvat i løbet af tilstande med nedsat kulhydrattilførsel indtræder i den hepatiske TCA-cyklus primært via anaplerose, dvs. ATP-afhængig carboxylation til oxaloacetat (OAA) eller til malat (MAL) og ikke oxidativ decarboxylering til acetyl-CoA (Jeoung et al., 2012; Magnusson et al., 1991; Merritt et al., 2011). I lever, bidrager glucose og pyruvat forsvindende med ketogenese, selv når pyruvatdecarboxylering til acetyl-CoA er maksimal (Jeoung et al., 2012).
Acetyl-CoA udgår adskillige roller integreret i metabolisme i leverformidlet ud over ATP-generationen via terminal oxidation (se også Integrationen af ketonkropsmetabolismen, posttranslationel modifikation og cellefysiologi). Acetyl-CoA aktiverer allosterisk (i) pyruvatcarboxylase (PC) og derved aktiverer en metabolisk kontrolmekanisme, som øger anaplerotisk indgang af metabolitter i TCA-cyklen (Owen et al., 2002, Scrutton and Utter, 1967) og (ii) pyruvatdehydrogenase kinase, som phosphorylerer og inhiberer pyruvatdehydrogenase (PDH) (Cooper et al., 1975), hvorved yderligere flow af pyruvat øges i TCA-cyklen via anaplerose. Endvidere katalyserer cytoplasmatisk acetyl-CoA, hvis pool forstærkes af mekanismer, der omdanner mitokondrialt acetyl-CoA til transportable metabolitter, hæmmer fedtsyreoxidation: acetyl-CoA carboxylase (ACC) katalyserer omdannelsen af acetyl-CoA til malonyl-CoA, det lipogene substrat og allosterisk hæmmer af mitokondrie CPT1 [gennemgået i (Kahn et al., 2005; McGarry og Foster, 1980)]. Således regulerer mitokondrieacetyl-CoA-puljen begge og reguleres af spildningsvejen for ketogenese, som orkestrerer nøgleaspekter af metabolisk metabolisme i leveren.
Ikke-Oxidative Metabolske Fates af Ketonlegemer
Den overvejende skæbne af leverafledte ketoner er SCOT-afhængig ekstrahepatisk oxidation. AcAc kan imidlertid eksporteres fra mitokondrier og anvendes i anabolske veje via omdannelse til AcAc-CoA ved en ATP-afhængig reaktion katalyseret af cytoplasmatisk acetoacetyl-CoA-syntetase (AACS, Fig. 1B). Denne vej er aktiv under hjernens udvikling og i lakterende brystkirtlen (Morris, 2005; Robinson og Williamson, 1978; Ohgami et al., 2003). AACS udtrykkes også stærkt i fedtvæv og aktiverede osteoklaster (Aguilo et al., 2010; Yamasaki et al., 2016). Cytoplasmisk AcAc-CoA kan enten styres af cytosolisk HMGCS1 mod sterolbiosyntese eller spaltes af enten to cytoplasmiske thiolaser til acetyl-CoA (ACAA1 og ACAT2), carboxyleret til malonyl-CoA og bidrager til syntese af fedtsyrer (Bergstrom et al., 1984; Edmond, 1974; Endemann et al., 1982; Geelen et al., 1983; Webber og Edmond, 1977).
Mens den fysiologiske betydning endnu ikke er fastslået, kan ketoner tjene som anabolske substrater selv i leveren. I kunstige eksperimentelle sammenhænge kan AcAc bidrage til så meget som halvdelen af nysyntetiseret lipid og op til 75% af nyt syntetiseret kolesterol (Endemann et al., 1982; Geelen et al., 1983; Freed et al., 1988). Fordi AcAc er afledt af ufuldstændig leverfedtoxidation, ville AcAc's evne til at bidrage til lipogenese in vivo indebære hepatisk forgæves cyklus, hvor fedtafledte ketoner kan bruges til lipidproduktion, en forestilling hvis fysiologiske betydning kræver eksperimentel validering, men kunne tjene adaptive eller utilpassede roller (Solinas et al., 2015). AcAc leverer ivrigt kolesterogenese med en lav AACS Km-AcAc (~50 �M), der favoriserer AcAc-aktivering selv i fødetilstand (Bergstrom et al., 1984). Den dynamiske rolle af cytoplasmatisk ketonmetabolisme er blevet foreslået i primære museembryonale neuroner og i 3T3-L1-afledte adipocytter, da AACS knockdown svækkede differentiering af hver celletype (Hasegawa et al., 2012a; Hasegawa et al., 2012b). Nedbrydning af AACS i mus in vivo reducerede serumkolesterol (Hasegawa et al., 2012c). SREBP-2, en master transkriptionel regulator af kolesterolbiosyntese og peroxisomproliferatoraktiveret receptor (PPAR)-? er AACS transkriptionelle aktivatorer og regulerer dets transkription under neuritudvikling og i leveren (Aguilo et al., 2010; Hasegawa et al., 2012c). Tilsammen kan cytoplasmatisk ketonlegememetabolisme være vigtig i udvalgte tilstande eller sygdomsnaturhistorier, men er utilstrækkelige til at skille sig af med leverafledte ketonstoffer, da massiv hyperketonemi forekommer i forbindelse med selektiv svækkelse af den primære oxidative skæbne via tab af funktionsmutationer til SCOT (Berry et al., 2001; Cotter et al., 2011).
Regulering af HMGCS2 og SCOT / OXCT1
Divergensen af et mitokondrium fra genet, der koder for cytosolisk HMGCS, forekom tidligt i udviklingen af hvirveldyr på grund af behovet for at understøtte leverketogenese hos arter med højere hjerne til kropsvægtforhold (Boukaftane et al., 1994, Cunnane og Crawford, 2003). Naturligt forekommende HMGCS2-mutationer i tab af funktion forårsager forekomster af hypoketotisk hypoglykæmi (Pitt et al., 2015; Thompson et al., 1997). Robust HMGCS2-ekspression er begrænset til hepatocytter og colonepitel, og dets ekspression og enzymatiske aktivitet koordineres gennem forskellige mekanismer (Mascaro et al., 1995; McGarry og Foster, 1980; Robinson og Williamson, 1980). Mens det fulde omfang af fysiologiske tilstande, der påvirker HMGCS2, kræver yderligere belysning, reguleres dets ekspression og / eller aktivitet i den tidlige postnatale periode, aldring, diabetes, sult eller indtagelse af ketogen kost (Balasse og Fery, 1989; Cahill GF Jr, 2006 ; Girard et al., 1992; Hegardt, 1999; Satapati et al., 2012; Sengupta et al., 2010). Hos fosteret korrelerer methylering af 5� flankerende region af Hmgcs2-genet omvendt med dets transkription og vendes delvist efter fødslen (Arias et al., 1995; Ayte et al., 1993; Ehara et al., 2015; Ferre et al. ., 1983). På samme måde udviser hepatisk Bdh1 et udviklingsmæssigt ekspressionsmønster, der stiger fra fødsel til fravænning, og induceres også ved ketogen diæt på en fibroblastvækstfaktor (FGF) -21-afhængig måde (Badman et al., 2007, Zhang et al., 1989 ). Ketogenese hos pattedyr er yderst lydhør over for både insulin og glucagon, henholdsvis undertrykt og stimuleret (McGarry og Foster, 1977). Insulin undertrykker fedtvævslipolyse og fratager således ketogenese af dets substrat, mens glucagon øger ketogen flux gennem en direkte virkning på leveren (Hegardt, 1999). Hmgcs2 transkription stimuleres af forkhead transkriptionsfaktor FOXA2, som hæmmes via insulin-phosphatidylinositol-3-kinase / Akt, og induceres ved glucagon-cAMP-p300-signalering (Arias et al., 1995; Hegardt, 1999; Quant et al. 1990; Thumelin et al., 1993; von Meyenn et al., 2013; Wolfrum et al., 2004; Wolfrum et al., 2003). PPAR? (Rodriguez et al., 1994) sammen med sit mål, FGF21 (Badman et al., 2007), inducerer også Hmgcs2-transkription i leveren under sult eller administration af ketogen diæt (Badman et al., 2007; Inagaki et al., 2007 ). Induktion af PPAR? kan forekomme før overgangen fra føtal til neonatal fysiologi, mens FGF21-aktivering kan foretrækkes i den tidlige neonatale periode via? OHB-medieret hæmning af histondeacetylase (HDAC) -3 (Rando et al., 2016). mTORC1 (pattedyrsmål for rapamycin kompleks 1) afhængig hæmning af PPAR? transkriptionsaktivitet er også en nøgleregulator for Hmgcs2-genekspression (Sengupta et al., 2010), og lever PER2, en mestersirkadisk oscillator, regulerer indirekte Hmgcs2-ekspression (Chavan et al., 2016). Nylige observationer indikerer, at ekstrahepatisk tumorinduceret interleukin-6 forringer ketogenese via PPAR? undertrykkelse (Flint et al., 2016).
HMGCS2 enzymaktivitet reguleres gennem flere PTM'er. HMGCS2 serinphosphorylering forøgede sin aktivitet in vitro (Grimsrud et al., 2012). HMGCS2-aktivitet inhiberes allosterisk af succinyl-CoA og lysinrestsuccinylering (Arias et al., 1995; Hegardt, 1999; Lowe og Tubbs, 1985; Quant et al., 1990; Rardin et al., 2013; Reed et al., 1975; Thumelin et al., 1993). Succinylering af HMGCS2-, HMGCL- og BDH1-lysinrester i levermitokondrier er mål for den NAD + -afhængige deacylase sirtuin 5 (SIRT5) (Rardin et al., 2013). HMGCS2-aktivitet forbedres også ved SIRT3-lysin-deacetylering, og det er muligt, at crosstalk mellem acetylering og succinylering regulerer HMGCS2-aktivitet (Rardin et al., 2013, Shimazu et al., 2013). På trods af disse PTMs evne til at regulere HMGCS2 Km og Vmax er svingninger af disse PTM'er endnu ikke blevet omhyggeligt kortlagt og er ikke blevet bekræftet som mekaniske drivkræfter for ketogenese in vivo.
SCOT udtrykkes i alle pattedyrceller, der bærer mitokondrier, undtagen de af hepatocytter. Vigtigheden af SCOT-aktivitet og ketolyse blev påvist i SCOT-KO-mus, som udviser ensartet dødelighed på grund af hyperketonemisk hypoglykæmi inden for 48h efter fødslen (Cotter et al., 2011). Vævsspecifik tab af SCOT i neuroner eller skelettmyocytter inducerer metaboliske abnormiteter under sult, men er ikke dødelig (Cotter et al., 2013b). Hos mennesker lever SCOT-mangel tidligt i livet med alvorlig ketoacidose, hvilket forårsager letargi, opkastning og koma (Berry et al., 2001; Fukao et al., 2000; Kassovska-Bratinova et al., 1996; Niezen-Koning et al. , 1997; Saudubray et al., 1987; Snyderman et al., 1998; Tildon og Cornblath, 1972). Forholdsvis lille er kendt på cellulær niveau omkring SCOT-gen og proteinekspressionsregulatorer. Oxct1 mRNA-ekspression og SCOT-protein og -aktivitet formindskes i ketotiske tilstande, muligvis gennem PPAR-afhængige mekanismer (Fenselau og Wallis, 1974, Fenselau og Wallis, 1976; Grinblat et al., 1986; Okuda et al., 1991; Turko et al. ., 2001; Wentz et al., 2010). Ved diabetisk ketoacidose bliver mismatchen mellem leverketogenese og ekstrahepatisk oxidation forværret ved nedsættelse af SCOT-aktivitet. Overekspression af insulinuafhængig glucosetransportør (GLUT1 / SLC2A1) i kardiomyocytter hæmmer også Oxct1-genekspression og nedregulerer ketones terminal oxidation i ikke-ketotisk tilstand (Yan et al., 2009). I leveren undertrykkes Oxct1 mRNA-overfladen af microRNA-122 og histon-methylering H3K27me3, som er tydelige under overgangen fra føtal til nyfødt periode (Thorrez et al., 2011). Imidlertid er undertrykkelse af hepatisk Oxct1-ekspression i postnatale perioden primært henført til evakueringen af Oxct1-udtrykkende hæmatopoietiske stamceller fra leveren, snarere end et tab af tidligere eksisterende Oxct1-ekspression i terminalt differentierede hepatocytter. Faktisk er ekspression af Oxct1 mRNA og SCOT protein i differentierede hepatocytter ekstremt lave (Orii et al., 2008).
SCOT er også reguleret af PTM'er. Enzymet er hyperacetyleret i hjerner fra SIRT3 KO-mus, som også udviser nedsat AcAc-afhængig acetyl-CoA-produktion (Dittenhafer-Reed et al., 2015). Ikke-enzymatisk nitrering af tyrosinrester af SCOT dæmper også dens aktivitet, hvilket er blevet rapporteret i hjerter fra forskellige diabetiske musemodeller (Marcondes et al., 2001; Turko et al., 2001; Wang et al., 2010a). I modsætning hertil øger nitrering af tryptofan-rester SCOT-aktivitet (Br�g�re et al., 2010; Rebrin et al., 2007). Molekylære mekanismer for restspecifik nitrering eller denitrering designet til at modulere SCOT-aktivitet kan eksistere og kræve belysning.
Kontroverser i ekstrahepatisk ketogenese
Hos pattedyr er det primære ketogene organ lever, og kun hepatocytter og tarmepitelceller udtrykker i rigelige mængder den mitokondrielle isoform af HMGCS2 (Cotter et al., 2013a; Cotter et al., 2014; McGarry og Foster, 1980; Robinson og Williamson, 1980) . Anaerob bakteriel fermentering af komplekse polysaccharider giver butyrat, som absorberes af colonocytter hos pattedyr til terminal oxidation eller ketogenese (Cherbuy et al., 1995), som kan spille en rolle i colonocytdifferentiering (Wang et al., 2016). Med undtagelse af tarmepitelceller og hepatocytter er HMGCS2 næsten fraværende i næsten alle andre pattedyrceller, men udsigten til ekstrahepatisk ketogenese er blevet øget i tumorceller, astrocytter i centralnervesystemet, nyrerne, bugspytkirtel ? celler, retinalt pigmentepitel (RPE) og endda i skeletmuskulatur (Adijanto et al., 2014; Avogaro et al., 1992; El Azzouny et al., 2016; Grabacka et al., 2016; Kang et al., 2015 ; Le Foll et al., 2014; Nonaka et al., 2016; Takagi et al., 2016a; Thevenet et al., 2016; Zhang et al., 2011). Ektopisk HMGCS2 er blevet observeret i væv, der mangler netto ketogen kapacitet (Cook et al., 2016; Wentz et al., 2010), og HMGCS2 udviser prospektive ketogenese-uafhængige "moonlighting"-aktiviteter, herunder inden for cellekernen (Chen et al. , 2016; Kostiuk et al., 2010; Meertens et al., 1998).
Ethvert ekstrahepatisk væv, der oxiderer ketonlegemer, har også potentialet til at akkumulere ketonlegemer via HMGCS2-uafhængige mekanismer (figur 2A). Der er dog ikke noget ekstrahepatisk væv, hvor en steady state ketonkropskoncentration overstiger den i cirkulationen (Cotter et al., 2011; Cotter et al., 2013b; Harrison og Long, 1940), hvilket understreger, at ketonlegemer transporteres ned ad en koncentrationsgradient via MCT1 / 2-afhængige mekanismer. En mekanisme for tilsyneladende ekstrahepatisk ketogenese kan faktisk afspejle relativ svækkelse af ketonoxidation. Yderligere potentielle forklaringer falder inden for dannelsen af ketonlegemer. For det første kan de novo ketogenese forekomme via reversibel enzymatisk aktivitet af thiolase og SCOT (Weidemann og Krebs, 1969). Når koncentrationen af acetyl-CoA er relativt høj, fungerer reaktioner, der normalt er ansvarlige for AcAc-oxidation i omvendt retning (GOLDMAN, 1954). En anden mekanisme opstår, når a-oxidationsafledte mellemprodukter akkumuleres på grund af en TCA-cyklusflaskehals, AcAc-CoA omdannes til l-? OHB-CoA gennem en reaktion katalyseret af mitokondrie 3-hydroxyacyl-CoA-dehydrogenase og yderligere ved 3-hydroxybutyryl CoA-deacylase til l-? OHB, som ikke kan skelnes ved massespektrometri eller resonansspektroskopi fra den fysiologiske enantiomer d-? OHB (Reed og Ozand, 1980). l-? OHB kan skelnes kromatografisk eller enzymatisk fra d-? OHB og er til stede i ekstrahepatisk væv, men ikke i lever eller blod (Hsu et al., 2011). Hepatisk ketogenese producerer kun d-? OHB, den eneste enantiomer, der er et BDH-substrat (Ito et al., 1984; Lincoln et al., 1987; Reed og Ozand, 1980; Scofield et al., 1982; Scofield et al., 1982). En tredje HMGCS2-uafhængig mekanisme genererer d-? OHB gennem aminosyrekatabolisme, især leucin og lysin. En fjerde mekanisme er kun synlig, fordi den skyldes en mærkningsgenstand og således betegnes som pseudoketogenese. Dette fænomen kan tilskrives reversibiliteten af SCOT- og thiolasereaktionerne og kan forårsage overvurdering af ketonlegemsomsætningen på grund af den isotopiske fortynding af ketonlegemsporeren i ekstrahepatisk væv (Des Rosiers et al., 1990; Fink et al., 1988) . Ikke desto mindre kan pseudoketogenese være ubetydelig i de fleste sammenhænge (Bailey et al., 1990; Keller et al., 1978). En skematisk skema (fig. 2A) indikerer en nyttig fremgangsmåde til anvendelse under overvejelse af forhøjet vævssteady-koncentration af ketoner.
� Nyre har for nylig fået opmærksomhed som et potentielt ketogent organ. I langt de fleste stater er nyren en nettoforbruger af leverafledte ketonstoffer, der udskiller eller reabsorberer ketonstoffer fra blodbanen, og nyrerne er generelt ikke en nettoketonlegemegenerator eller -koncentrator (Robinson og Williamson, 1980). Forfatterne af en klassisk undersøgelse konkluderede, at minimal renal ketogenese kvantificeret i et kunstigt eksperimentelt system ikke var fysiologisk relevant (Weidemann og Krebs, 1969). For nylig er nyreketogenese blevet udledt i diabetiske og autofagi-deficiente musemodeller, men det er mere sandsynligt, at multiorganskift i metabolisk homeostase ændrer integrativ ketonmetabolisme gennem input på flere organer (Takagi et al., 2016a; Takagi et al., 2016b; Zhang et al., 2011). En nylig publikation foreslog renal ketogenese som en beskyttende mekanisme mod iskæmi-reperfusionsskade i nyren (Tran et al., 2016). Absolutte steady state-koncentrationer af ?OHB fra ekstrakter af musenyrevæv blev rapporteret ved ~4~12 mM. For at teste, om dette var holdbart, kvantificerede vi ?OHB-koncentrationer i nyreekstrakter fra fodrede og 24 timers fastende mus. Serum-OHB-koncentrationer steg fra ~100 �M til 2 mM med 24 timers faste (fig. 2B), mens steady-state OHB-koncentrationer i nyrerne ca. 100 �M i fødetilstand og kun 1 mM i 24 timers fastende tilstand (fig. 2C�E), observationer, der er i overensstemmelse med koncentrationer kvantificeret for over 45 år siden (Hems og Brosnan, 1970). Det er fortsat muligt, at leverafledte ketonlegemer i ketotiske tilstande kan være genbeskyttende, men beviser for renal ketogenese kræver yderligere underbyggelse. Overbevisende beviser, der understøtter ægte ekstrahepatisk ketogenese, blev præsenteret i RPE (Adijanto et al., 2014). Denne spændende metaboliske transformation blev foreslået for potentielt at tillade RPE-afledte ketoner at strømme til fotoreceptor- eller M�ller glia-celler, hvilket kunne hjælpe med regenereringen af fotoreceptorens ydre segment.
? OHB som signalformidler
Selvom de er energirigt rige, udøver ketonlegemer provokerende "ikke-kanoniske" signaleringsroller i cellulær homeostase (Fig. 3) (Newman og Verdin, 2014; Rojas-Morales et al., 2016). For eksempel hæmmer ?OHB Klasse I HDAC'er, hvilket øger histonacetylering og derved inducerer ekspressionen af gener, der begrænser oxidativt stress (Shimazu et al., 2013). ?OHB i sig selv er en histon kovalent modifikator på lysinrester i leveren af fastende eller streptozotocin-inducerede diabetiske mus (Xie et al., 2016) (se også nedenfor, Integration af ketonlegememetabolisme, post-translationel modifikation og cellefysiologi, og Ketonlegemer, oxidativt stress og neurobeskyttelse).
�
?OHB er også en effektor via G-protein koblede receptorer. Gennem uklare molekylære mekanismer undertrykker det sympatisk nervesystems aktivitet og reducerer det samlede energiforbrug og hjertefrekvens ved at hæmme kortkædet fedtsyresignalering gennem G-proteinkoblet receptor 41 (GPR41) (Kimura et al., 2011). En af de mest undersøgte signaleffekter af ?OHB fortsætter gennem GPR109A (også kendt som HCAR2), et medlem af hydrocarboxylsyre-GPCR-underfamilien udtrykt i fedtvæv (hvidt og brunt) (Tunaru et al., 2003) og i immunceller (Ahmed et al., 2009). ?OHB er den eneste kendte endogene ligand af GPR109A-receptoren (EC50 ~770 �M) aktiveret af d-?OHB, l-?OHB og butyrat, men ikke AcAc (Taggart et al., 2005). Den høje koncentrationstærskel for GPR109A-aktivering opnås gennem overholdelse af en ketogen diæt, sult eller under ketoacidose, hvilket fører til hæmning af fedtvævslipolyse. Den anti-lipolytiske virkning af GPR109A fortsætter gennem inhibering af adenylylcyclase og nedsat cAMP, hvilket hæmmer hormonfølsom triglyceridlipase (Ahmed et al., 2009; Tunaru et al., 2003). Dette skaber en negativ feedback-loop, hvor ketose sætter en modulerende bremse på ketogenesen ved at mindske frigivelsen af ikke-esterificerede fedtsyrer fra adipocytter (Ahmed et al., 2009; Taggart et al., 2005), en effekt der kan opvejes af den sympatiske drift, der stimulerer lipolysen. Niacin (vitamin B3, nikotinsyre) er en potent (EC50 ~ 0.1 �M) ligand for GRP109A, som er effektivt anvendt i årtier mod dyslipidæmi (Benyo et al., 2005; Benyo et al., 2006; Fabbrini et al., 2010a; Lukasova et al., 2011; Tunaru et al., 2003). Mens niacin øger omvendt kolesteroltransport i makrofager og reducerer aterosklerotiske læsioner (Lukasova et al., 2011), forbliver virkningerne af ?OHB på aterosklerotiske læsioner ukendte. Selvom GPR109A-receptoren udøver beskyttende roller, og der eksisterer spændende forbindelser mellem brug af ketogen diæt i slagtilfælde og neurodegenerative sygdomme (Fu et al., 2015; Rahman et al., 2014), er en beskyttende rolle for ?OHB via GPR109A ikke blevet påvist in vivo .
Endelig kan OHB påvirke appetit og mæthed. En meta-analyse af undersøgelser, der målte virkningerne af ketogene diæter og diæter med meget lav energi, konkluderede, at deltagere, der indtager disse diæter, udviser højere mæthed sammenlignet med kontroldiæter (Gibson et al., 2015). Men en plausibel forklaring på denne effekt er de yderligere metaboliske eller hormonelle elementer, der kan modulere appetitten. For eksempel udviste mus, der blev holdt på en gnaver ketogen diæt, øget energiforbrug sammenlignet med foderkontrol-fodrede mus, på trods af lignende kalorieindtag, og cirkulerende leptin eller gener af peptider, der regulerer fodringsadfærd, blev ikke ændret (Kennedy et al., 2007). Blandt de foreslåede mekanismer, der tyder på appetitundertrykkelse af ?OHB, omfatter både signalering og oxidation (Laeger et al., 2010). Hepatocytspecifik deletion af døgnrytme-genet (Per2) og kromatin-immunpræcipitationsundersøgelser afslørede, at PER2 direkte aktiverer Cpt1a-genet og indirekte regulerer Hmgcs2, hvilket fører til svækket ketose i Per2 knockout-mus (Chavan et al., 2016). Disse mus udviste svækket madforventning, som delvist blev genoprettet ved systemisk ?OHB-administration. Fremtidige undersøgelser vil være nødvendige for at bekræfte centralnervesystemet som et direkte ?OHB-mål, og om ketonoxidation er nødvendig for de observerede effekter, eller om en anden signalmekanisme er involveret. Andre efterforskere har påberåbt sig muligheden for lokal astrocyt-afledt ketogenese i den ventromediale hypothalamus som en regulator af fødeindtagelse, men disse foreløbige observationer vil også drage fordel af genetiske og flux-baserede vurderinger (Le Foll et al., 2014). Forholdet mellem ketose og næringsstofmangel er fortsat interessant, fordi sult og mæthed er vigtige elementer i mislykkede vægttabsforsøg.
Integration af Ketonlegemetabolisme, Post-Translational Modifikation, og Cell Physiology
Ketonlegemer bidrager til compartmentaliserede pools af acetyl-CoA, et nøglemellemprodukt, der udviser fremtrædende roller i cellulær metabolisme (Pietrocola et al., 2015). En rolle acetyl-CoA er at tjene som et substrat for acetylering, en enzymatisk katalyseret histonkovalent modifikation (Choudhary et al., 2014; Dutta et al., 2016; Fan et al., 2015; Menzies et al., 2016 ). Et stort antal dynamisk acetylerede mitochondriale proteiner, hvoraf mange kan forekomme gennem ikke-enzymatiske mekanismer, har også fremkommet fra beregningsproteomikstudier (Dittenhafer-Reed et al., 2015; Hebert et al., 2013; Rardin et al., 2013 Shimazu et al., 2010). Lysin-deacetylaser bruger en zinkkofaktor (fx nukleocytosoliske HDAC'er) eller NAD + som co-substrat (sirtuiner, SIRT'er) (Choudhary et al., 2014; Menzies et al., 2016). Acetylproteomet tjener som både sensor og effektor af den totale cellulære acetyl-CoA-pool, da fysiologiske og genetiske manipulationer hver især resulterer i ikke-enzymatiske globale variationer af acetylering (Weinert et al., 2014). Da intracellulære metabolitter tjener som modulatorer for acetylering af lysinrest, er det vigtigt at overveje ketonorganernes rolle, hvis overflade er yderst dynamisk.
OHB er en epigenetisk modifikator gennem mindst to mekanismer. Øgede OHB-niveauer induceret ved faste, kaloribegrænsning, direkte administration eller langvarig træning fremkalder HDAC-hæmning eller histonacetyltransferase-aktivering (Marosi et al., 2016; Sleiman et al., 2016) eller til oxidativ stress (Shimazu et al., 2013) . OHB-hæmning af HDAC3 kunne regulere nyfødt metabolisk fysiologi (Rando et al., 2016). Uafhængigt ændrer? OHB selv histonlysinrester direkte (Xie et al., 2016). Langvarig faste eller steptozotocin-induceret diabetisk ketoacidose øgede histon-a-hydroxybutyrylering. Selvom antallet af lysin-a-hydroxybutyrylering- og acetyleringssteder var sammenlignelige, blev der observeret støkiometrisk større histon-a-hydroxybutyrylering end acetylering. Tydelige gener blev påvirket af histonlysin-a-hydroxybutyrylering versus acetylering eller methylering, hvilket antydede forskellige cellulære funktioner. Om a-hydroxybutyrylering er spontan eller enzymatisk vides ikke, men udvider rækkevidden af mekanismer gennem ketonlegemer, der dynamisk påvirker transkription.
Væsentlige celleomprogrammeringshændelser under kaloribegrænsning og næringsstofnedsættelse kan formidles i henholdsvis SIRT3- og SIRT5-afhængig mitokondrie-deacetylering og desuccinylering, hvilket regulerer ketogene og ketolytiske proteiner på posttranslationelt niveau i lever- og ekstrahepatisk væv (Dittenhafer-Reed et al., 2015; Hebert et al., 2013; Rardin et al., 2013; Shimazu et al., 2010). Selvom støkiometrisk sammenligning af besatte steder ikke nødvendigvis knytter sig direkte til forskydninger i metabolisk flux, er mitokondrieacetylering dynamisk og kan drives af acetyl-CoA-koncentration eller mitokondrie-pH snarere end enzymatiske acetyltransferaser (Wagner og Payne, 2013). At SIRT3 og SIRT5 modulerer aktiviteter af ketonlegemer, der metaboliserer enzymer, fremkalder spørgsmålet om ketonernes gensidige rolle i skulptur af acetylproteom, succinylproteom og andre dynamiske cellulære mål. Da variationer af ketogenese afspejler NAD + -koncentrationer, kunne ketonproduktion og overflod faktisk regulere sirtuinaktivitet og derved påvirke total acetyl-CoA / succinyl-CoA-puljer, acylproteomet og dermed mitokondrie- og cellefysiologi. a-hydroxybutyrylering af enzymlysinrester kunne tilføje endnu et lag til cellulær omprogrammering. I ekstrahepatisk væv kan ketonlegemeoxidation stimulere analoge ændringer i cellehomeostase. Mens opdeling af acetyl-CoA-puljer er stærkt reguleret og koordinerer et bredt spektrum af cellulære ændringer, kræver ketonlegemers evne til direkte at forme både mitokondrie- og cytoplasmatiske acetyl-CoA-koncentrationer belysning (Chen et al., 2012; Corbet et al., 2016; Pougovkina et al., 2014; Schwer et al., 2009; Wellen og Thompson, 2012). Fordi acetyl-CoA-koncentrationer er tæt reguleret, og acetyl-CoA er membranimpermeant, er det afgørende at overveje drivmekanismerne, der koordinerer acetyl-CoA-homeostase, herunder produktionshastigheder og terminal oxidation i TCA-cyklussen, omdannelse til ketonlegemer, mitokondrie udstrømning via carnitinacetyltransferase (CrAT) eller acetyl-CoA-eksport til cytosol efter omdannelse til citrat og frigivelse ved ATP-citratlyase (ACLY). Nøglerollerne for disse sidstnævnte mekanismer i celleacetylproteom og homeostase kræver matchet forståelse af rollerne for ketogenese og ketonoxidation (Das et al., 2015; McDonnell et al., 2016; Moussaieff et al., 2015; Overmyer et al., 2015; Seiler et al., 2014; Seiler et al., 2015; Wellen et al., 2009; Wellen and Thompson, 2012). Konvergente teknologier inden for metabolomics og acylproteomics i indstillingen af genetisk manipulerede modeller vil være forpligtet til at specificere mål og resultater.
Anti- og pro-inflammatoriske reaktioner på ketonorganer
Ketose og ketonlegemer modulerer inflammation og immuncellefunktion, men forskellige og endda uoverensstemmende mekanismer er blevet foreslået. Langvarig mangel på næringsstoffer reducerer betændelse (Youm et al., 2015), men den kroniske ketose af type 1-diabetes er en pro-inflammatorisk tilstand (Jain et al., 2002; Kanikarla-Marie og Jain, 2015; Kurepa et al., 2012 ). Mekanisme-baserede signalroller for? OHB i inflammation opstår, fordi mange immunsystemceller, herunder makrofager eller monocytter, i høj grad udtrykker GPR109A. Mens? OHB udviser et overvejende antiinflammatorisk respons (Fu et al., 2014; Gambhir et al., 2012; Rahman et al., 2014; Youm et al., 2015), kan høje koncentrationer af ketonlegemer, især AcAc, muligvis udløse et proinflammatorisk respons (Jain et al., 2002; Kanikarla-Marie og Jain, 2015; Kurepa et al., 2012).
Anti-inflammatoriske roller af GPR109A-ligander i åreforkalkning, fedme, inflammatorisk tarmsygdom, neurologisk sygdom og kræft er blevet gennemgået (Graff et al., 2016). GPR109A-ekspression forstærkes i RPE-celler fra diabetiske modeller, humane diabetespatienter (Gambhir et al., 2012) og i mikroglia under neurodegeneration (Fu et al., 2014). Antiinflammatoriske virkninger af? OHB forstærkes af GPR109A-overekspression i RPE-celler og ophæves ved farmakologisk hæmning eller genetisk knockout af GPR109A (Gambhir et al., 2012). OHB og eksogen nikotinsyre (Taggart et al., 2005), begge giver antiinflammatoriske virkninger i TNF? eller LPS-induceret betændelse ved at nedsætte niveauerne af proinflammatoriske proteiner (iNOS, COX-2) eller secernerede cytokiner (TNFa, IL-1a, IL-6, CCL2 / MCP-1), delvist gennem inhibering af NF -? B-translokation (Fu et al., 2014; Gambhir et al., 2012). ? OHB nedsætter ER-stress og NLRP3-inflammasomet og aktiverer det antioxidative stressrespons (Bae et al., 2016; Youm et al., 2015). I neurodegenerativ inflammation involverer GPR109A-afhængig? OHB-medieret beskyttelse imidlertid ikke inflammatoriske mediatorer som MAPK pathway-signalering (f.eks. ERK, JNK, p38) (Fu et al., 2014), men kan kræve COX-1-afhængig PGD2 produktion (Rahman et al., 2014). Det er spændende, at makrofag GPR109A kræves for at udøve en neurobeskyttende effekt i en iskæmisk slagmodel (Rahman et al., 2014), men evnen hos? OHB til at hæmme NLRP3-inflammatoren i knoglemarvafledte makrofager er GPR109A uafhængig (Youm et al. ., 2015). Selvom de fleste undersøgelser forbinder? OHB med antiinflammatoriske virkninger, kan? OHB være pro-inflammatorisk og øge markører for lipidperoxidering i kalvehepatocytter (Shi et al., 2014). Anti- versus proinflammatoriske virkninger af? OHB kan således afhænge af celletype,? OHB-koncentration, eksponeringsvarighed og tilstedeværelsen eller fraværet af co-modulatorer.
I modsætning til? OHB kan AcAc aktivere proinflammatorisk signalering. Forhøjet AcAc, især med en høj glukosekoncentration, intensiverer endotelcelleskader gennem en NADPH oxidase / oxidativ stressafhængig mekanisme (Kanikarla-Marie og Jain, 2015). Høje AcAc-koncentrationer i navlestrengen hos diabetiske mødre var korreleret med højere proteinoxidationshastighed og MCP-1-koncentration (Kurepa et al., 2012). Høj AcAc hos diabetespatienter var korreleret med TNF? ekspression (Jain et al., 2002) og AcAc, men ikke? OHB, induceret TNF ?, MCP-1-ekspression, ROS-akkumulering og nedsat cAMP-niveau i U937 humane monocytceller (Jain et al., 2002; Kurepa et al. ., 2012).
Ketonkropsafhængige signalfænomener udløses ofte kun med høje ketonkropskoncentrationer (> 5 mM), og i tilfælde af mange undersøgelser, der forbinder ketoner til pro- eller antiinflammatoriske virkninger gennem uklare mekanismer. Desuden, på grund af de modstridende virkninger af? OHB versus AcAc på inflammation, og evnen af AcAc /? OHB-forhold til at påvirke mitokondrie redox potentiale, sammenligner de bedste eksperimenter, der vurderer ketonlegemers roller på cellulære fænotyper, virkningerne af AcAc og? OHB i forskellige forhold og ved varierende kumulative koncentrationer [fx (Saito et al., 2016)]. Endelig kan AcAc kun købes kommercielt som et lithiumsalt eller som en ethylester, der kræver basehydrolyse inden brug. Lithiumkation inducerer uafhængigt signaltransduktionskaskader (Manji et al., 1995), og AcAc-anion er labil. Endelig kan undersøgelser, der anvender racemisk d / l-? OHB, forveksles, da kun d-? OHB-stereoisomeren kan oxideres til AcAc, men d-? OHB og l-? OHB kan hver signalere gennem GPR109A, hæmme NLRP3-inflammatoren, og tjener som lipogene substrater.
Ketonlegemer, Oxidativ Stress og Neuroprotektion
Oxidativ stress er typisk defineret som en tilstand, hvor ROS præsenteres i overskud på grund af overdreven produktion og/eller nedsat eliminering. Antioxidant- og oxidativ stress-dæmpende roller af ketonlegemer er blevet beskrevet bredt både in vitro og in vivo, især i sammenhæng med neurobeskyttelse. Da de fleste neuroner ikke effektivt genererer højenergifosfater fra fedtsyrer - men oxiderer ketonstoffer, når der er mangel på kulhydrater, er neurobeskyttende virkninger af ketonstoffer særligt vigtige (Cahill GF Jr, 2006; Edmond et al., 1987; Yang et al., 1987). I oxidativ stressmodeller tyder BDH1-induktion og SCOT-undertrykkelse på, at ketonlegememetabolisme kan omprogrammeres til at opretholde forskellig cellesignalering, redoxpotentiale eller metaboliske krav (Nagao et al., 2016; Tieu et al., 2003).
Ketonlegemer reducerer graden af cellulær skade, skade, død og lavere apoptose i neuroner og kardiomyocytter (Haces et al., 2008; Maalouf et al., 2007; Nagao et al., 2016; Tieu et al., 2003). Påberåbte mekanismer er varierede og ikke altid lineært relateret til koncentration. Lave millimolære koncentrationer af (d eller l)-?OHB fjerner ROS (hydroxylanion), mens AcAc fjerner adskillige ROS-arter, men kun ved koncentrationer, der overstiger det fysiologiske område (IC50 20-67 mM) (Haces et al., 2008) . Omvendt er en gavnlig indflydelse på elektrontransportkædens redoxpotentiale en mekanisme, der almindeligvis er forbundet med d-OHB. Mens alle tre ketonlegemer (d/l-?OHB og AcAc) reducerede neuronal celledød og ROS-akkumulering udløst af kemisk hæmning af glykolyse, forhindrede kun d-?OHB og AcAc neuronal ATP-nedgang. Omvendt, i en hypoglykæmisk in vivo-model forhindrede (d eller l)-?OHB, men ikke AcAc hippocampus lipidperoxidation (Haces et al., 2008; Maalouf et al., 2007; Marosi et al., 2016; Murphy, 2009 ; Tieu et al., 2003). In vivo undersøgelser af mus fodret med en ketogen diæt (87 % kcal fedt og 13 % protein) udviste neuroanatomisk variation af antioxidantkapacitet (Ziegler et al., 2003), hvor de mest dybtgående ændringer blev observeret i hippocampus med stigning i glutathionperoxidase og total antioxidant kapacitet.
Ketogen diæt, ketonestere (se også Terapeutisk brug af ketogen diæt og eksogene ketonlegemer) eller ?OHB-administration udøver neurobeskyttelse i modeller for iskæmisk slagtilfælde (Rahman et al., 2014); Parkinsons sygdom (Tieu et al., 2003); ilttoksicitetsanfald i centralnervesystemet (D'Agostino et al., 2013); epileptiske spasmer (Yum et al., 2015); mitokondriel encefalomyopati, laktatacidose og slagtilfælde-lignende (MELAS) episoder syndrom (Frey et al., 2016) og Alzheimers sygdom (Cunnane og Crawford, 2003; Yin et al., 2016). Omvendt påviste en nylig rapport histopatologiske beviser for neurodegenerativ progression ved en ketogen diæt i en transgen musemodel af unormal mitokondriel DNA-reparation, på trods af stigninger i mitokondriel biogenese og antioxidantsignaturer (Lauritzen et al., 2016). Andre modstridende rapporter tyder på, at eksponering for høje ketonlegemekoncentrationer fremkalder oxidativt stress. Høje ?OHB- eller AcAc-doser inducerede nitrogenoxidsekretion, lipidperoxidation, reduceret ekspression af SOD, glutathionperoxidase og katalase i kalve-hepatocytter, mens MAPK-pathway-induktionen i rotte-hepatocytter blev tilskrevet AcAc, men ikke ?OHB (Abdelmegeed et al., 2004. ; Shi et al., 2014; Shi et al., 2016).
Samlet set forbinder de fleste rapporter? OHB til dæmpning af oxidativ stress, da administrationen hæmmer ROS / superoxidproduktion, forhindrer lipidperoxidering og proteinoxidation, øger antioxidantproteinniveauer og forbedrer mitokondrie respiration og ATP produktion (Abdelmegeed et al., 2004; Haces et al., 2008; Jain et al., 1998; Jain et al., 2002; Kanikarla-Marie and Jain, 2015; Maalouf et al., 2007; Maalouf and Rho, 2008; Marosi et al., 2016; Tieu et al., 2003; Yin et al., 2016; Ziegler et al., 2003). Mens AcAc er blevet mere direkte korreleret end? OHB med induktion af oxidativt stress, dissekeres disse effekter ikke altid let fra potentielle proinflammatoriske reaktioner (Jain et al., 2002; Kanikarla-Marie og Jain, 2015; Kanikarla-Marie og Jain, 2016). Desuden er det kritisk at overveje, at den tilsyneladende antioxidative fordel, der tilvejebringes af pleiotropiske ketogene diæter, muligvis ikke transduceres af ketonlegemer i sig selv, og neurobeskyttelse, der tildeles af ketonlegemer, muligvis ikke helt kan tilskrives oxidativ stress. For eksempel under glukosemangel, i en model af glukosmangel i kortikale neuroner, stimulerede? OHB autofagisk flux og forhindrede akkumulering af autophagosom, som var forbundet med nedsat neuronal død (Camberos-Luna et al., 2016). d-? OHB inducerer også de kanoniske antioxidantproteiner FOXO3a, SOD, MnSOD og katalase, prospektivt gennem HDAC-inhibering (Nagao et al., 2016; Shimazu et al., 2013).
Ikke-alkoholisk fedtsyge-sygdom (NAFLD) og ketonlegemetabolisme
Fedme-associeret NAFLD og ikke-alkoholisk steatohepatitis (NASH) er de mest almindelige årsager til leversygdom i vestlige lande (Rinella og Sanyal, 2016), og NASH-induceret leversvigt er en af de mest almindelige årsager til levertransplantation. Mens overskydende opbevaring af triacylglyceroler i hepatocytter> 5% af levervægt (NAFL) alene ikke forårsager degenerativ leverfunktion, korrelerer progressionen til NAFLD hos mennesker med systemisk insulinresistens og øget risiko for type 2-diabetes og kan bidrage til patogenesen af hjerte-kar-sygdom og kronisk nyresygdom (Fabbrini et al., 2009; Targher et al., 2010; Targher og Byrne, 2013). De patogene mekanismer i NAFLD og NASH er ufuldstændigt forstået, men inkluderer abnormiteter i hepatocytmetabolisme, hepatocyt-autofagi og endoplasmatisk retikulumstress, leverimmun cellefunktion, fedtvævbetændelse og systemiske inflammatoriske mediatorer (Fabbrini et al., 2009; Masuoka og Chalasani, 2013 ; Targher et al., 2010; Yang et al., 2010). Forstyrrelser af kulhydrat-, lipid- og aminosyremetabolisme forekommer i og bidrager til fedme, diabetes og NAFLD hos mennesker og i modelorganismer [gennemgået i (Farese et al., 2012; Lin og Accili, 2011; Newgard, 2012; Samuel og Shulman, 2012; Sun og Lazar, 2013)]. Mens hepatocytabnormiteter i cytoplasmatisk lipidmetabolisme ofte observeres i NAFLD (Fabbrini et al., 2010b), er rollen som mitokondrie-metabolisme, som styrer oxidativ bortskaffelse af fedt, mindre klar i NAFLD-patogenesen. Abnormaliteter af mitokondrie metabolisme forekommer i og bidrager til NAFLD / NASH patogenese (Hyotylainen et al., 2016; Serviddio et al., 2011; Serviddio et al., 2008; Wei et al., 2008). Der er generelle (Felig et al., 1974; Iozzo et al., 2010; Koliaki et al., 2015; Satapati et al., 2015; Satapati et al., 2012; Sunny et al., 2011) men ikke ensartede ( Koliaki og Roden, 2013; Perry et al., 2016; Rector et al., 2010) enighed om, at før udviklingen af bona fide NASH, lever mitokondrieoxidation og især fedtoxidation øges i fedme, systemisk insulinresistens og NAFLD. Det er sandsynligt, at efterhånden som NAFLD skrider frem, kommer oxidativ kapacitet heterogenitet, selv blandt individuelle mitokondrier, og i sidste ende bliver oxidativ funktion svækket (Koliaki et al., 2015; Rector et al., 2010; Satapati et al., 2008; Satapati et al. ., 2012).
Ketogenese bruges ofte som en proxy for hepatisk fedtoxidation. Forringelser af ketogenese dukker op, efterhånden som NAFLD skrider frem i dyremodeller og sandsynligvis hos mennesker. Gennem ufuldstændigt definerede mekanismer undertrykker hyperinsulinemi ketogenese, hvilket muligvis bidrager til hypoketonemi sammenlignet med magre kontroller (Bergman et al., 2007; Bickerton et al., 2008; Satapati et al., 2012; Soeters et al., Sunny et al., 2009; , 2011; Vice et al., 2005). Ikke desto mindre er evnen til at cirkulere ketonlegemekoncentrationer til at forudsige NAFLD kontroversiel (M�nnist� et al., 2015; Sanyal et al., 2001). Robuste kvantitative magnetiske resonansspektroskopiske metoder i dyremodeller afslørede øget ketonomsætningshastighed med moderat insulinresistens, men faldende hastigheder var tydelige med mere alvorlig insulinresistens (Satapati et al., 2012; Sunny et al., 2010). Hos overvægtige mennesker med fedtlever er den ketogene hastighed normal (Bickerton et al., 2008; Sunny et al., 2011), og derfor er ketogenesehastigheden formindsket i forhold til den øgede fedtsyrebelastning i hepatocytter. Som følge heraf kan a-oxidationsafledt acetyl-CoA være rettet mod terminal oxidation i TCA-cyklussen, hvilket øger terminal oxidation, phosphoenolpyruvat-drevet gluconeogenese via anaplerose/kataplerose og oxidativt stress. Acetyl-CoA undergår muligvis også eksport fra mitokondrier som citrat, et forløbersubstrat for lipogenese (Fig. 4) (Satapati et al., 2015; Satapati et al., 2012; Solinas et al., 2015). Mens ketogenese bliver mindre følsom over for insulin eller faste med langvarig fedme (Satapati et al., 2012), forbliver de underliggende mekanismer og nedstrøms konsekvenser af dette ufuldstændigt forstået. Nylige beviser indikerer, at mTORC1 undertrykker ketogenese på en måde, der kan være nedstrøms for insulinsignalering (Kucejova et al., 2016), hvilket er i overensstemmelse med observationerne om, at mTORC1 hæmmer PPAR?-medieret Hmgcs2-induktion (Sengupta et al., 2010) ( se også Regulering af HMGCS2 og SCOT/OXCT1).
�
Foreløbige observationer fra vores gruppe tyder på negative leverkonsekvenser af ketogen insufficiens (Cotter et al., 2014). For at teste hypotesen om, at svækket ketogenese, selv i kulhydratfyldte og dermed "ikke-ketogene" tilstande, bidrager til unormal glukosemetabolisme og fremkalder steatohepatitis, genererede vi en musemodel af markant ketogen insufficiens ved administration af antisense oligonukleotider målrettede (AS) Hmgcs2. Tab af HMGCS2 i almindelige, fedtfattige chow-fodrede voksne mus forårsagede mild hyperglykæmi og markant øget produktion af hundredvis af levermetabolitter, hvoraf en række kraftigt antydede aktivering af lipogenese. Fodring med højt fedtindhold af mus med utilstrækkelig ketogenese resulterede i omfattende hepatocytskade og betændelse. Disse resultater understøtter de centrale hypoteser om, at (i) ketogenese ikke er en passiv overløbsvej, men snarere en dynamisk knude i hepatisk og integreret fysiologisk homeostase, og (ii) forsigtig ketogen forstærkning for at afbøde NAFLD/NASH og forstyrret leverglukosemetabolisme er værd at udforske .
Hvordan kan nedsat ketogenese bidrage til leverskade og ændret glukosehomeostase? Den første overvejelse er, om synderen er mangel på ketogen flux eller ketoner i sig selv. En nylig rapport antyder, at ketonlegemer kan afbøde oxidativ stressinduceret leverskade som reaktion på n-3 flerumættede fedtsyrer (Pawlak et al., 2015). Husk på, at ketonlegemer ikke oxideres på grund af mangel på SCOT-ekspression i hepatocytter, men de kan bidrage til lipogenese og tjene en række signaleringsroller uafhængigt af deres oxidation (se også ikke-oxidative metaboliske skæbne til ketonlegemer og? OHB som en signalformidler). Det er også muligt, at ketonlegemer, der er afledt af hepatocytter, kan tjene som et signal og / eller metabolit for nærliggende celletyper i den hepatiske acinus, herunder stjerneceller og Kupffer-cellemakrofager. Mens den begrænsede tilgængelige litteratur antyder, at makrofager ikke er i stand til at oxidere ketonlegemer, er dette kun blevet målt ved hjælp af klassiske metoder og kun i peritoneale makrofager (Newsholme et al., 1986; Newsholme et al., 1987), hvilket indikerer, at en vurdering er passende givet rigelig SCOT-ekspression i knoglemarvafledte makrofager (Youm et al., 2015).
Hepatocyt ketogen flux kan også være cytobeskyttende. Mens de sundhedsmæssige mekanismer muligvis ikke afhænger af ketogenese i sig selv, er ketogene diæter med lavt kulhydratindhold blevet forbundet med forbedring af NAFLD (Browning et al., 2011; Foster et al., 2010; Kani et al., 2014; Schugar og Crawford, 2012) . Vores observationer indikerer, at hepatocyt-ketogenese kan give feedback og regulere TCA-cyklusflux, anaplerotisk flux, phosphoenolpyruvat-afledt gluconeogenese (Cotter et al., 2014) og endda glykogenomsætning. Ketogen svækkelse leder acetyl-CoA til at øge TCA-flux, som i leveren er blevet forbundet med øget ROS-medieret skade (Satapati et al., 2015; Satapati et al., 2012); tvinger omledning af kulstof til de novo syntetiserede lipidarter, der kunne vise sig at være cytotoksiske; og forhindrer NADH re-oxidation til NAD+ (Cotter et al., 2014) (fig. 4). Samlet set er fremtidige eksperimenter nødvendige for at adressere mekanismer, hvorigennem relativ ketogen insufficiens kan blive maladaptiv, bidrage til hyperglykæmi, fremkalde steatohepatitis, og om disse mekanismer er operante i human NAFLD/NASH. Da epidemiologiske beviser tyder på nedsat ketogenese under progressionen af steatohepatitis (Embade et al., 2016; Marinou et al., 2011; M�nnist� et al., 2015; Pramfalk et al., 2015; Safaei et al.), 2016. terapier, der øger hepatisk ketogenese, kan vise sig at være gavnlige (Degirolamo et al., 2016; Honda et al., 2016).
Ketonlegemer og hjertesvigt (HF)
Med en metabolisk hastighed på over 400 kcal/kg/dag og en omsætning på 6-35 kg ATP/dag er hjertet det organ med det højeste energiforbrug og det højeste oxidative behov (Ashrafian et al., 2007; Wang et al., 2010b). Langt størstedelen af myokardiets energiomsætning ligger inden for mitokondrier, og 70% af denne forsyning stammer fra FAO. Hjertet er altædende og fleksibelt under normale forhold, men det patologisk omdannende hjerte (f.eks. på grund af hypertension eller myokardieinfarkt) og det diabetiske hjerte bliver hver især metabolisk ufleksible (Balasse og Fery, 1989; BING, 1954; Fukao et al., 2004 ; Lopaschuk et al., 2010; Taegtmeyer et al., 1980; Taegtmeyer et al., 2002; Young et al., 2002). Faktisk fremkalder genetisk programmerede abnormiteter af hjertebrændstofmetabolisme i musemodeller kardiomyopati (Carley et al., 2014; Neubauer, 2007). Under fysiologiske forhold oxiderer normale hjerter ketonlegemer i forhold til deres levering på bekostning af fedtsyre- og glucoseoxidation, og myokardium er den højeste ketonlegemeforbruger pr. masseenhed (BING, 1954; Crawford et al., 2009; GARLAND et al. ., 1962; Hasselbaink et al., 2003; Jeffrey et al., 1995; Pelletier et al., 2007; Tardif et al., 2001; Yan et al., 2009). Sammenlignet med fedtsyreoxidation er ketonstoffer mere energieffektive, hvilket giver mere energi tilgængelig til ATP-syntese pr. molekyle investeret ilt (P/O-forhold) (Kashiwaya et al., 2010; Sato et al., 1995; Veech, 2004) . Ketonlegemeoxidation giver også potentielt højere energi end FAO, hvilket holder ubiquinon oxideret, hvilket øger redoxspændvidden i elektrontransportkæden og gør mere energi tilgængelig til at syntetisere ATP (Sato et al., 1995; Veech, 2004). Oxidation af ketonstoffer kan også begrænse ROS-produktionen og dermed oxidativt stress (Veech, 2004).
Preliminære interventions- og observationsstudier indikerer en potentiel velgørende rolle af ketonlegemer i hjertet. I den eksperimentelle iskæmi / reperfusionsskadesammensætning tillod ketonlegemer potentielle kardioprotektive virkninger (Al-Zaid et al., 2007; Wang et al., 2008), muligvis på grund af øget mitokondriell overflod i hjertet eller opregulering af afgørende oxidativ phosphorylering mediatorer (Snorek et al., 2012; Zou et al., 2002). Nylige undersøgelser indikerer, at ketonkroppens udnyttelse er forøget i manglende hjerter hos mus (Aubert et al., 2016) og mennesker (Bedi et al., 2016), der støtter forudgående observationer hos mennesker (BING, 1954; Fukao et al., 2000; Janardhan et al., 2011; Longo et al., 2004; Rudolph og Schinz, 1973; Tildon og Cornblath, 1972). Kropskoncentrationer i cirkulerende keton øges hos patienter med hjertesvigt, i direkte forhold til påfyldningstryk, observationer hvis mekanisme og betydning forbliver ukendt (Kupari et al., 1995; Lommi et al., 1996; Lommi et al., 1997; Neely et al. ., 1972), men mus med selektiv SCOT-mangel i cardiomyocytter udviser accelereret patologisk ventrikulær remodeling og ROS-signaturer som svar på kirurgisk induceret trykoverbelastningsskade (Schugar et al., 2014).
Nyligt spændende observationer i diabetesbehandling har vist en potentiel sammenhæng mellem myokardisk ketonmetabolisme og patologisk ventrikulær remodeling (figur 5). Inhibering af den renale proximale tubulære natrium / glucose-co-transportør 2 (SGLT2i) øger koncentrationen af ketonkroppen i mennesker (Ferrannini et al., 2016a; Inagaki et al., 2015) og mus (Suzuki et al., 2014) via øget hepatisk ketogenese (Ferrannini et al., 2014, Ferrannini et al., 2016a; Katz og Leiter, 2015; Mudaliar et al., 2015). Påfaldende faldt mindst en af disse stoffer HF-hospitalisering (fx som afsløret af EMPA-REG OUTCOME-forsøg) og forbedret kardiovaskulær dødelighed (Fitchett et al., 2016; Sonesson et al., 2016; Wu et al., 2016a Zinman et al., 2015). Selvom førermekanismerne bag positive HF-resultater til forbundne SGLT2i forbliver aktivt debatteret, er overlevelsesfordelen sandsynligvis multifaktorial, fremadrettet inklusiv ketosis, men også salutære virkninger på vægt-, blodtryk-, glukose- og urinsyre niveauer, arteriel stivhed, sympatisk nervesystem, osmotisk diurese / reduceret plasmavolumen og øget hæmatokrit (Raz og Cahn, 2016, Vallon og Thomson, 2016). Tværtimod forbliver tanken om, at terapeutisk stigende ketonæmi enten hos HF-patienter eller dem med høj risiko for at udvikle HF, kontroversiel, men er under aktiv undersøgelse i prækliniske og kliniske studier (Ferrannini et al., 2016b; Kolwicz et al., 2016; Lopaschuk og Verma, 2016; Mudaliar et al., 2016; Taegtmeyer, 2016).
�
Ketonlegemer i kræftbiologi
Forbindelser mellem ketonlegemer og kræft udvikler sig hurtigt, men studier i både dyremodeller og mennesker har givet forskellige konklusioner. Fordi ketonmetabolisme er dynamisk og næringsstoftilstand, er det lokkende at forfølge biologiske forbindelser til kræft på grund af muligheden for præcisionsstyrede ernæringsmæssige terapier. Kræftceller undergår metabolisk omprogrammering for at opretholde hurtig celleproliferation og vækst (DeNicola og Cantley, 2015; Pavlova og Thompson, 2016). Den klassiske Warburg-effekt i kræftcellemetabolismen stammer fra den dominerende rolle glykolyse og mælkesyrefermentation for at overføre energi og kompensere for lavere afhængighed af oxidativ phosphorylering og begrænset mitokondriell respiration (De Feyter et al., 2016; Grabacka et al., 2016; Kang et al., 2015; Poff et al., 2014; Shukla et al., 2014). Glucosecarbon er primært rettet gennem glycolyse, pentosephosphatvejen og lipogenese, som tilsammen tilvejebringer mellemprodukter, der er nødvendige for ekspansion af tumorbiomasse (Grabacka et al., 2016; Shukla et al., 2014; Yoshii et al., 2015). Tilpasning af kræftceller til glukoseafrivning sker gennem evnen til at udnytte alternative brændstofkilder, herunder acetat, glutamin og aspartat (Jaworski et al., 2016; Sullivan et al., 2015). For eksempel afslører begrænset adgang til pyruvat cancercellernes evne til at omdanne glutamin til acetyl-CoA ved carboxylering og opretholde både energetiske og anabolske behov (Yang et al., 2014). En interessant tilpasning af cancerceller er anvendelsen af acetat som et brændstof (Comerford et al., 2014; Jaworski et al., 2016; Mashimo et al., 2014; Wright og Simone, 2016; Yoshii et al., 2015). Acetat er også et substrat for lipogenese, hvilket er kritisk for tumorcelleproliferation, og forstærkning af denne lipogene kanal er forbundet med kortere patientoverlevelse og større tumorbyrde (Comerford et al., 2014; Mashimo et al., 2014; Yoshii et al. ., 2015).
Ikke-kræftceller skifter nemt deres energikilde fra glukose til ketonlegemer under glukosemangel. Denne plasticitet kan være mere variabel blandt kræftcelletyper, men in vivo implanterede hjernetumorer oxiderede [2,4-13C2]-?OHB i samme grad som omgivende hjernevæv (De Feyter et al., 2016). �Omvendt Warburg-effekt� eller �to-kompartment tumormetabolisme�-modeller antager, at cancerceller inducerer OHB-produktion i tilstødende fibroblaster, der leverer tumorcellens energibehov (Bonuccelli et al., 2010; Martinez-Outschoorn et al.), 2012. . I leveren er et skift i hepatocytter fra ketogenese til ketonoxidation i hepatocellulære carcinomceller (hepatom) i overensstemmelse med aktivering af BDH1- og SCOT-aktiviteter observeret i to hepatomcellelinjer (Zhang et al., 1989). Faktisk udtrykker hepatomceller OXCT1 og BDH1 og oxiderer ketoner, men kun når serum sultede (Huang et al., 2016). Alternativt er tumorcelleketogenese også blevet foreslået. Dynamiske skift i ketogen genekspression er udstillet under cancerøs transformation af tyktarmsepitel, en celletype, der normalt udtrykker HMGCS2, og en nylig rapport antydede, at HMGCS2 kan være en prognostisk markør for dårlig prognose i kolorektale og pladecellekarcinomer (Camarero et al., 2006; Chen et al., 2016). Hvorvidt denne forening kræver eller involverer ketogenese eller en måneskinsfunktion af HMGCS2, skal stadig afgøres. Omvendt, tilsyneladende ?OHB-produktion af melanom- og glioblastomceller, stimuleret af PPAR? agonist fenofibrat, var forbundet med vækststop (Grabacka et al., 2016). Yderligere undersøgelser er nødvendige for at karakterisere rollerne af HMGCS2/SCOT-ekspression, ketogenese og ketonoxidation i cancerceller.
Ud over brændstofmetabolismens område er ketoner for nylig blevet impliceret i kræftcellebiologi via en signalmekanisme. Analyse af BRAF-V600E + melanom indikerede OCT1-afhængig induktion af HMGCL på en onkogen BRAF-afhængig måde (Kang et al., 2015). HMGCL-forstærkning blev korreleret med højere cellulær AcAc-koncentration, hvilket igen forbedrede BRAFV600E-MEK1-interaktion, hvilket forstærkede MEK-ERK-signalering i en feed-forward-loop, der driver tumorcelleproliferation og -vækst. Disse observationer rejser det spændende spørgsmål om potentiel ekstrahepatisk ketogenese, der derefter understøtter en signalmekanisme (se også OHB som en signalmægler og kontroverser i ekstrahepatisk ketogenese). Det er også vigtigt at overveje uafhængige virkninger af AcAc, d-? OHB og l-? OHB på kræftmetabolisme, og når man overvejer HMGCL, kan leucinkatabolisme også blive forstyrret.
Effekterne af ketogene diæter (se også Terapeutisk brug af ketogen diæt og eksogene ketonstoffer) i cancerdyremodeller er varierede (De Feyter et al., 2016; Klement et al., 2016; Meidenbauer et al., 2015; Poff et al. ., 2014; Seyfried et al., 2011; Shukla et al., 2014). Mens epidemiologiske sammenhænge mellem fedme, kræft og ketogene diæter diskuteres (Liskiewicz et al., 2016; Wright og Simone, 2016), antydede en metaanalyse, der anvender ketogene diæter i dyremodeller og i menneskelige undersøgelser, en gavnlig indvirkning på overlevelse, med fordele prospektivt forbundet med størrelsen af ketose, tidspunktet for diætstart og tumorplacering (Klement et al., 2016; Woolf et al., 2016). Behandling af bugspytkirtelkræftceller med ketonstoffer (d-?OHB eller AcAc) hæmmede vækst, proliferation og glykolyse, og en ketogen diæt (81 % kcal fedt, 18 % protein, 1 % kulhydrat) reducerede in vivo tumorvægt, glykæmi og øget muskel- og kropsvægt hos dyr med implanteret cancer (Shukla et al., 2014). Lignende resultater blev observeret ved brug af en metastatisk glioblastomcellemodel i mus, der modtog ketontilskud i kosten (Poff et al., 2014). Omvendt øgede en ketogen diæt (91 % kcal fedt, 9 % protein) den cirkulerende "OHB-koncentration og formindsket glykæmi", men havde ingen indflydelse på hverken tumorvolumen eller overlevelsesvarighed hos gliombærende rotter (De Feyter et al., 2016). Et glukoseketonindeks er blevet foreslået som en klinisk indikator, der forbedrer metabolisk styring af ketogen diæt-induceret hjernekræftbehandling hos mennesker og mus (Meidenbauer et al., 2015). Samlet set er rollerne af ketonlegememetabolisme og ketonlegemer i cancerbiologi fristende, fordi de hver især udgør håndterbare terapeutiske muligheder, men grundlæggende aspekter mangler at blive belyst, med klare påvirkninger, der kommer fra en matrix af variabler, herunder (i) forskelle mellem eksogen keton legemer versus ketogen diæt, (ii) cancercelletype, genomiske polymorfismer, grad og stadie; og (iii) timing og varighed af eksponering for den ketotiske tilstand.
Ketogenese skabes af ketonlegemer gennem nedbrydning af fedtsyrer og ketogene aminosyrer. Denne biokemiske proces giver energi til forskellige organer, specielt hjernen, under faste forhold som et svar på en utilgængelighed af blodglukose. Ketonlegemer produceres hovedsageligt i mitokondrier af leverceller. Mens andre celler er i stand til at udføre ketogenese, er de ikke så effektive til at gøre det som leverceller. Fordi ketogenese forekommer i mitokondrier, reguleres dets processer uafhængigt. Dr. Alex Jimenez DC, CCST Insight
Terapeutisk anvendelse af Ketogenic Diet og Exogenous Ketone Bodies
Anvendelsen af ketogen diæt og ketonstoffer som terapeutiske redskaber er også opstået i ikke-cancerøse sammenhænge, herunder fedme og NAFLD/NASH (Browning et al., 2011; Foster et al., 2010; Schugar og Crawford, 2012); hjertesvigt (Huynh, 2016; Kolwicz et al., 2016; Taegtmeyer, 2016); neurologisk og neurodegenerativ sygdom (Martin et al., 2016; McNally og Hartman, 2012; Rho, 2015; Rogawski et al., 2016; Yang og Cheng, 2010; Yao et al., 2011); medfødte metabolismefejl (Scholl-Burgi et al, 2015); og træningspræstation (Cox et al., 2016). Effektiviteten af ketogen diæt er især blevet værdsat i terapi af epileptiske anfald, især hos lægemiddelresistente patienter. De fleste undersøgelser har evalueret ketogen diæt hos pædiatriske patienter og afslører op til en ~50% reduktion i anfaldshyppighed efter 3 måneder, med forbedret effektivitet i udvalgte syndromer (Wu et al., 2016b). Erfaringen er mere begrænset i voksen epilepsi, men en lignende reduktion er tydelig, med bedre respons hos symptomatisk generaliseret epilepsi patienter (Nei et al., 2014). De underliggende anti-konvulsive mekanismer forbliver uklare, selvom postulerede hypoteser omfatter reduceret glukoseudnyttelse/glykolyse, omprogrammeret glutamattransport, indirekte påvirkning af ATP-følsom kaliumkanal eller adenosin A1-receptor, ændring af natriumkanalisoform-ekspression eller effekter på cirkulerende hormoner inklusive leptin ( Lambrechts et al., 2016; Lin et al., 2017; Lutas og Yellen, 2013). Det er fortsat uklart, om den anti-konvulsive effekt primært kan tilskrives ketonstoffer eller på grund af de kaskade metaboliske konsekvenser af diæter med lavt kulhydratindhold. Ikke desto mindre synes ketonestere (se nedenfor) at hæve anfaldstærsklen i dyremodeller af fremkaldte anfald (Ciarlone et al., 2016; D'Agostino et al., 2013; Viggiano et al., 2015).
Atkins-stil og ketogen, lavt kulhydrat kostvaner anses ofte for ubehagelige og kan forårsage forstoppelse, hyperuricæmi, hypokalcæmi, hypomagnesæmi, føre til nefrolithiasis, ketoacidose, forårsage hyperglykæmi og øge koncentrationen af cirkulerende cholesterol og fri fedtsyre (Bisschop et al., 2001 Kossoff og Hartman, 2012; Kwiterovich et al., 2003; Suzuki et al., 2002). Af disse årsager udgør langsigtet adherence udfordringer. Gnagereundersøgelser bruger almindeligvis en særskilt makronæringsfordeling (94% kcalfedt, 1% kcal carbohydrat, 5% kcal protein, Bio-Serv F3666), hvilket fremkalder en robust ketose. Imidlertid øger proteinindholdet selv til 10% kcal væsentligt ketosis, og 5% kcal proteinrestriktion tillader konfronterende metaboliske og fysiologiske virkninger. Denne diætformulering er også cholinudarmet, en anden variabel, som påvirker følsomheden over for leverskader og endog ketogenese (Garbow et al., 2011; Jornayvaz et al., 2010; Kennedy et al., 2007; Pissios et al., 2013; Schugar et al., 2013). Virkninger af langvarigt forbrug af ketogene diæter hos mus forbliver ufuldstændigt defineret, men nyere undersøgelser hos mus afslørede normal overlevelse og fraværet af leverskademarkører hos mus på ketogene diæter i løbet af deres levetid, selv om aminosyremetabolisme, energiforbrug og insulinsignalering blev markant omprogrammeret (Douris et al., 2015).
Mekanismer, der øger ketosis gennem mekanismer, der er alternative til ketogene kostvaner, indbefatter anvendelsen af indtagelige ketonlegemer. Administrering af eksogene ketonlegemer kan skabe en unik fysiologisk tilstand, der ikke opstår i normal fysiologi, fordi cirkulerende glukose- og insulinkoncentrationer er relativt normale, mens cellerne kan spare glucoseoptagelse og -udnyttelse. Ketonlegemer har selv korte halveringstider, og indtagelse eller infusion af natrium-OHB-salt for at opnå terapeutisk ketose fremkalder en uheldig natriumbelastning. R / S-1,3-butandiol er en ikke-toksisk dialkohol, der let oxideres i leveren til opnåelse af d / l- OHB (Desrochers et al., 1992). I forskellige eksperimentelle sammenhænge er denne dosis administreret dagligt til mus eller rotter i så længe som syv uger, hvilket giver cirkulerende? OHB-koncentrationer på op til 5 mM inden for 2 timer efter administration, hvilket er stabilt i mindst yderligere 3 timer (D ' Agostino et al., 2013). Delvis undertrykkelse af fødeindtaget er blevet observeret hos gnavere givet R / S-1,3-butandiol (Carpenter og Grossman, 1983). Derudover er tre kemisk adskilte ketonestere (KE'er), (i) monoester af R-1,3-butandiol og d-aOHB (R-3-hydroxybutylR-? OHB); (ii) glyceryl-tris-? OHB; og (iii) R, S-1,3-butandiolacetoacetat-diester, er også blevet grundigt undersøgt (Brunengraber, 1997; Clarke et al., 2012a; Clarke et al., 2012b; Desrochers et al., 1995a; Desrochers et al. ., 1995b; Kashiwaya et al., 2010). En iboende fordel ved førstnævnte er, at der produceres 2 mol fysiologisk d-? OHB pr. Mol KE efter esterasehydrolyse i tarmen eller leveren. Sikkerhed, farmakokinetik og tolerance er blevet undersøgt mest omfattende hos mennesker, der indtager R-3-hydroxybutyl R-? OHB, ved doser op til 714 mg / kg, hvilket giver cirkulerende d-? OHB-koncentrationer op til 6 mM (Clarke et al., 2012a; Cox et al., 2016; Kemper et al., 2015; Shivva et al., 2016). Hos gnavere nedsætter denne KE kalorindtag og plasma totalt kolesterol, stimulerer brun fedtvæv og forbedrer insulinresistens (Kashiwaya et al., 2010, Kemper et al., 2015; Veech, 2013). Nylige fund indikerer, at R-3-hydroxybutyl R-? OHB-indtagelse under træning hos trænede atleter nedsatte skeletmuskelglykolyse og plasmalactatkoncentrationer, øget intramuskulær triacylglyceroloxidation og bevaret muskelglykogenindhold, selv når co-indtaget kulhydrat stimuleret insulinsekretion ( Cox et al., 2016). Yderligere udvikling af disse spændende resultater er påkrævet, fordi forbedringen i udholdenhedsøvelsen var overvejende drevet af et robust svar på KE i 2 / 8-emnerne. Ikke desto mindre understøtter disse resultater klassiske undersøgelser, som indikerer en præference for ketonoxidation over andre substrater (GARLAND et al., 1962; Hasselbaink et al., 2003; Stanley et al., 2003; Valente-Silva et al., 2015), herunder under træning, og at uddannede atleter kan være mere primede til at udnytte ketoner (Johnson et al., 1969a; Johnson og Walton, 1972; Winder et al., 1974; Winder et al., 1975). Endelig må de mekanismer, der understøtter forbedret træningsevne efter samme kalorieindtag (differentielt fordelt mellem makronæringsstoffer) og lige iltforbrugstal, bestemmes.
Fremtidsperspektiv
Når først i høj grad var stigmatiseret som en overløbsvej, der er i stand til at akkumulere toksiske emissioner fra fedtforbrænding i kulhydratbegrænsede tilstande (det ketotoksiske paradigme), understøtter nyere observationer forestillingen om, at ketonlegememetabolisme tjener en gavnlig rolle selv i kulhydratfyldte tilstande, hvilket åbner en ketohormetik � hypotese. Mens de lette ernæringsmæssige og farmakologiske tilgange til at manipulere ketonmetabolisme gør det til et attraktivt terapeutisk mål, forbliver aggressivt stillede, men forsigtige eksperimenter i både de grundlæggende og translationelle forskningslaboratorier. Uopfyldte behov er opstået inden for områderne for at definere rollen af at udnytte ketonmetabolisme i hjertesvigt, fedme, NAFLD/NASH, type 2-diabetes og cancer. Omfanget og virkningen af ketonlegemers 'ikke-kanoniske' signaleringsroller, herunder regulering af PTM'er, der sandsynligvis feeds tilbage og fremad i metaboliske og signalveje, kræver dybere udforskning. Endelig kunne ekstrahepatisk ketogenese åbne spændende parakrine og autokrine signalmekanismer og muligheder for at påvirke co-metabolisme i nervesystemet og tumorer for at opnå terapeutiske mål.
Som konklusion er ketonstoffer skabt af leveren for at blive brugt som energikilde, når der ikke er nok glukose let tilgængeligt i menneskekroppen. Ketogenese opstår, når der er lave glukoseniveauer i blodet, især efter at andre cellulære kulhydratlagre er opbrugt. Formålet med artiklen ovenfor var at diskutere de multidimensionelle roller af ketonlegemer i brændstofmetabolisme, signalering og terapi. Omfanget af vores information er begrænset til kiropraktik og rygsøjlesundhedsproblemer. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på�915-850-0900 .
Rygsmerte er en af de mest udbredte årsager til handicap og ubesvarede arbejdsdage over hele verden. Rygsmerter tilskrives den næst hyppigste årsag til lægehjælp, der kun overtales af infektioner i øvre luftveje. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Rygsøjlen er en kompleks struktur, der består af knogler, led, ledbånd og muskler, blandt andet blødt væv. Skader og / eller forværrede forhold, såsom herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller bilulykkesskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe med at lindre rygsmerter gennem brug af spinaljusteringer og manuelle manipulationer, hvilket i sidste ende forbedrer smertelindring. �
nukleær erythroid 2-relateret faktor 2 signalveje, bedst kendt som Nrf2, er en beskyttelsesmekanisme, der fungerer som en "master regulator" af menneskekroppens antioxidant respons. Nrf2 registrerer niveauerne af oxidativ stress i cellerne og udløser beskyttende antioxidantmekanismer. Mens Nrf2 aktivering kan have mange fordele, kan Nrf2 "overekspression" have flere risici.
Det ser ud til, at en afbalanceret grad af NRF2 er afgørende for at forhindre den samlede udvikling af en række sygdomme ud over den generelle forbedring af disse sundhedsmæssige problemer. Imidlertid kan NRF2 også forårsage komplikationer. Hovedårsagen til NRF2 "overekspression" skyldes en genetisk mutation eller en fortsat kronisk eksponering for et kemisk eller oxidativt stress blandt andre. Nedenfor vil vi diskutere nedskæringerne af Nrf2 overekspression og demonstrere dets virkningsmekanismer i den menneskelige krop.
Kræft
Forskningsundersøgelser viste, at mus, der ikke udtrykker NRF2, er mere tilbøjelige til at udvikle kræft som reaktion på fysisk og kemisk stimulering. Lignende forskningsundersøgelser viste imidlertid, at NRF2-overaktivering eller endda KEAP1-inaktivering kan resultere i forværring af visse kræftformer, især hvis disse veje er blevet afbrudt. Overaktiv NRF2 kan forekomme ved rygning, hvor kontinuerlig NRF2-aktivering menes at være årsagen til lungekræft hos rygere. Nrf2-overekspression kan medføre, at kræftceller ikke selvdestrueres, mens intermitterende NRF2-aktivering kan forhindre kræftceller i at udløse toksininduktion.
Fordi NRF2-overekspression øger den menneskelige krops antioxidantevne til at fungere ud over redox-homeostase, øger dette celledeling og genererer et unaturligt mønster af DNA- og histon-methylering. Dette kan i sidste ende gøre�kemoterapi og strålebehandling mindre effektiv mod kræft. Derfor kan begrænsning af NRF2-aktivering med stoffer som DIM, Luteolin, Zi Cao eller salinomycin være ideel for patienter med cancer, selvom Nrf2-overaktivering ikke bør anses for at være den eneste årsag til cancer. Næringsmangler kan påvirke gener, herunder NRF2. Dette kan være en måde at se, hvordan mangler bidrager til tumorer.
Lever
Overaktivering af Nrf2 kan også påvirke funktionen af specifikke organer i menneskekroppen. NRF2 overekspression kan i sidste ende blokere produktionen af den insulinlignende vækstfaktor 1, eller IGF-1, fra leveren, som er afgørende for regenerering af leveren.
Hjerte
Selvom den akutte overekspression af Nrf2 kan have sine fordele, kan kontinuerlig overekspression af NRF2 forårsage langsigtede skadelige virkninger på hjertet, såsom kardiomyopati. NRF2-ekspression kan forøges gennem høje niveauer af cholesterol eller aktiveringen af HO-1. Dette menes at være årsagen til, at kroniske forhøjede niveauer af kolesterol kan forårsage kardiovaskulære sundhedsproblemer.
Vitiligo
NRF2-overekspression er også blevet påvist at hæmme evnen til at repigere i vitiligo, da det kan forhindre tyrosinase- eller TYR-virkning, som er essentiel for repigmentering gennem melaninogenese. Forskningsundersøgelser har vist, at denne proces kan være en af de primære årsager til, hvorfor mennesker med vitiligo ikke synes at aktivere Nrf2 så effektivt som mennesker uden vitiligo.
Hvorfor NRF2 ikke fungerer korrekt
hormesis
NRF2 skal aktiveres hormetisk for at kunne drage fordel af dets fordele. Med andre ord bør Nrf2 ikke udløse hvert minut eller hver dag,�derfor er det en god idé at tage pauser fra det, for eksempel 5 dage på 5 fridage eller hver anden dag. NRF2 skal også opnå en specifik tærskel for at udløse dens hormetiske respons, hvor en lille stressfaktor måske ikke er nok til at udløse den.
DJ-1 Oxidation
Protein deglycase DJ-1, eller bare DJ-1, også kaldet Parkinsons sygdomsprotein eller PARK7, er en master regulator og detektor af redoxstatus i menneskekroppen. DJ-1 er afgørende for at regulere, hvor længe NRF2 kan udføre sin funktion og producere et antioxidantrespons. I tilfælde af at DJ-1 bliver overoxideret, vil cellerne gøre DJ-1 proteinet mindre tilgængeligt.
Denne proces inducerer NRF2-aktivering til at udløbe for hurtigt, da DJ-1 er afgørende for at opretholde balancerede niveauer af NRF2 og forhindre dem i at blive brudt ned i cellen. Hvis DJ-1-proteinet ikke eksisterer eller overoxideres, vil NRF2-ekspression sandsynligvis være minimal, selv ved anvendelse af DIM eller alternative NRF2-aktivatorer. DJ-1-udtryk er afgørende for at genskabe forringet NRF2-handling.
Kronisk sygdom
Hvis du har en kronisk sygdom, inklusive CIRS, kroniske infektioner/dysbiose/SIBO eller tungmetalopbygning, såsom kviksølv og/eller det fra rodkanaler, kan disse blokere systemerne for NRF2 og fase to-afgiftning. I stedet for at oxidativ stress forvandler NRF2 til en antioxidant, vil NRF2 ikke udløse, og oxidativ stress kan forblive i cellen og forårsage skade, hvilket betyder, at der ikke er nogen antioxidantrespons. Dette er en væsentlig grund til, at mange mennesker med CIRS har adskillige følsomheder og når til adskillige faktorer. Nogle mennesker tror, de kan have en herx-reaktion, men denne reaktion kan kun beskadige cellerne længere.
Behandling af kronisk sygdom vil dog tillade leveren at udlede toksiner i galden, og gradvist udvikle den hormetiske respons af NRF2-aktivering. Hvis galden forbliver giftig, og den ikke udskilles fra den menneskelige krop, vil den genaktivere NRF2's oxidative stress og få dig til at føle dig værre, når den først er reabsorberet fra mave-tarmkanalen. For eksempel kan ochratoxin A blokere NRF2. Udover at behandle problemet kan histon-deacetylase-hæmmere blokere den oxidative reaktion fra en række af de faktorer, der udløser NRF2-aktivering, men det kan også forhindre NRF2 i at trigge normalt, hvilket måske i sidste ende ikke opfylder sit formål.
Fish Oil Dysregulation
Cholinergics er stoffer, der hæver acetylcholin, eller ACh, og cholin i hjernen gennem forøgelsen af ACh, især når det hæmmer nedbrydningen af ACh. Patienter med CIRS har ofte problemer med dysreguleringen af acetylcholinniveauer i menneskekroppen, især i hjernen. Fiskolier udløser NRF2, der aktiverer sin beskyttende antioxidantmekanisme i cellerne.
Mennesker med kroniske sygdomme kan have problemer med kognitiv stress og acetylcholin excitotoksicitet, fra ophobning af organofosfat, som kan få fiskeolie til at skabe betændelse i menneskekroppen. Cholinmangel inducerer desuden NRF2-aktivering. At inkludere cholin i din kost (polyfenoler, æg osv.) kan hjælpe med at forstærke virkningerne af kolinerg dysregulering.
Hvad reducerer NRF2?
Faldende NRF2 overekspression er bedst for mennesker, der har kræft, selv om det kan være gavnligt for en række andre sundhedsmæssige problemer.
Kost, kosttilskud og almindelige lægemidler:
Apigenin (højere doser)
Brucea javanica
Kastanier
EGCG (høje doser øger NRF2)
Fenugreek (Trigonelline)
Hiba (Hinokitiol /? -Thujaplicin)
High Salt Diet
Luteolin (selleri, grøn peber, persille, perilla blade og kamille te - højere doser kan øge NRF2 - 40 mg / kg luteolin tre gange om ugen)
Metformin (kronisk indtagelse)
N-acetyl-L-cystein (NAC, ved blokering af det oxidative respons, især ved høje doser)
Orange Peel (har polymethoxylerede flavonoider)
Quercetin (højere doser kan øge NRF2 - 50 mg / kg / d quercetin)
Salinomycin (lægemiddel)
Retinol (all-trans retinsyre)
C-vitamin kombineret med Quercetin
Zi Cao (Lilla Gromwel har Shikonin / Alkannin)
Stier og Andet:
Bach1
BET
biofilm
Brusatol
camptothecin
DNMT
DPP-23
EZH2
Glucocorticoid Receptor signalering (Dexamethason og Betamethason også)
GSK-3? (feedback fra lovgivningen)
HDAC aktivering?
halofuginon
Homocystein (ALCAR kan reversere denne homocystein inducerer lave niveauer af NRF2)
IL-24
Keap1
MDA-7
NF? B
Ochratoxin A (aspergillus og pencicllium arter)
Promyelocytisk leukæmiprotein
p38
p53
p97
Retinsyre-receptor-alfa
Selenite
SYVN1 (Hrd1)
STAT3-hæmning (såsom kryptotanshinon)
Testosteron (og testosteron propionat, selv om TP intranasalt kan øge NRF2)
Trecator (Ethionamid)
Trx1 (via reduktion af Cys151 i Keap1 eller Cys506 i NLS-regionen af Nrf2)
Trolox
Vorinostat
Zinkmangel (gør det værre i hjernen)
Nrf2 Handlingsmekanisme
Oxidativ stress udløser gennem CUL3, hvor NRF2 fra KEAP1, en negativ hæmmer, kommer senere i kernerne i disse celler, stimulerer transkriptionen af ARE'erne, omdanner sulfider til disulfider og omdanner dem til flere antioxidantgener, hvilket fører til opregulering af antioxidanter, såsom som GSH, GPX, GST, SOD osv. Resten af disse kan ses i listen nedenfor:
Forøger AKR
Øger ARE
Forøger ATF4
Forøger Bcl-xL
Forøger Bcl-2
Øger BDNF
Forøger BRCA1
Øger c-juni
Forøger CAT
Forøger cGMP
Forøger CKIP-1
Forøger CYP450
Forøger Cul3
Forøger GCL
Forøger GCLC
Forøger GCLM
Forøger GCS
Forøger GPx
Forøger GR
Forøger GSH
Forøger GST
Forøger HIF1
Forøger HO-1
Øger HQO1
Forøger HSP70
Forøger IL-4
Forøger IL-5
Forøger IL-10
Forøger IL-13
Forøger K6
Forøger K16
Forøger K17
Forøger mEH
Forhøjer Mrp2-5
Forøger NADPH
Øger hak 1
Øger NQO1
Forøger PPAR-alpha
Øger Prx
Forøger p62
Øger SesN2
Forøger Slco1b2
Øger sMafs
Øger SOD
Øger Trx
Øger Txn (d)
Øger UGT1 (A1 / 6)
Forøger VEGF
Reducerer ADAMTS (4 / 5)
Reducerer alfa-SMA
Reducerer ALT
Reducerer AP1
Reducerer AST
Reducerer Bach1
Reducerer COX-2
Reducerer DNMT
Reducerer FASN
Reducerer FGF
Reducerer HDAC
Reducerer IFN-?
Reducerer IgE
Reducerer IGF-1
Reducerer IL-1b
Reducerer IL-2
Reducerer IL-6
Reducerer IL-8
Reducerer IL-25
Reducerer IL-33
Reducerer iNOS
Reducerer LT
Reducerer Keap1
Reducerer MCP-1
Reducerer MIP-2
Reducerer MMP-1
Reducerer MMP-2
Reducerer MMP-3
Reducerer MMP-9
Reducerer MMP-13
Reducerer NfkB
Reducerer NO
Reducerer SIRT1
Reducerer TGF-b1
Reducerer TNF-alpha
Reducerer Tyr
Reducerer VCAM-1
Kodet fra NFE2L2-genet, NRF2 eller nukleær erythroid 2-relateret faktor 2, er en transkriptionsfaktor i den grundlæggende leucin-lynlås, eller bZIP, superfamilien, der anvender en Cap'n'Collar- eller CNC-struktur.
Det fremmer nitrogeniske enzymer, biotransformationsenzymer og xenobiotiske effluxtransportører.
Det er en vigtig regulator ved induktionen af fase II-antioxidant- og afgiftningsenzymgener, som beskytter celler mod skader forårsaget af oxidativt stress og elektrofile angreb.
Under homøostatiske tilstande sekventeres Nrf2 i cytosolen via kropslig binding af det N-terminale domæne af Nrf2 eller det Kelch-lignende ECH-associerede protein eller Keap1, også omtalt som INrf2 eller inhibitor af Nrf2, hæmning af Nrf2-aktivering.
Det kan også kontrolleres af pattedyr selenoprotein thioredoxinreduktase 1 eller TrxR1, som fungerer som en negativ regulator.
Ved sårbarhed overfor elektrofile stressorer dissocierer Nrf2 fra Keap1, der translokerer ind i kernen, hvor den derefter heterodimeriseres med et område af transkriptionel regulatorisk protein.
Hyppige interaktioner omfatter dem fra transkriptionsmyndighederne Jun og Fos, som kan være medlemmer af aktivatorproteinfamilien af transkriptionsfaktorer.
Efter dimerisering binder disse komplekser derefter til antioxidant / elektrofile responsive komponenter ARE / EpRE og aktiverer transkription, som det er tilfældet med Jun-Nrf2-komplekset eller undertrykker transkription, ligesom Fos-Nrf2-komplekset.
Placeringen af ARE, som udløses eller hæmmes, vil bestemme, hvilke gener der kontrolleres transkriptionelt af disse variabler.
Når ARE udløses:
Aktivering af syntesen af antioxidanter er i stand til at afgifte ROS som katalase, superoxid-dismutase eller SOD, GSH-peroxidaser, GSH-reduktase, GSH-transferase, NADPH-quinonoxidoreduktase eller NQO1, Cytochrom P450 monooxygenase-system, thioredoxigenase-system, thioredoxigenase reduktase og HSP70.
Aktivering af denne GSH-syntase tillader en mærkbar vækst af den intracellulære GSH-grad, hvilket er ret beskyttende.
Forøgelsen af denne syntese og grader af fase II enzymer som UDP-glucuronosyltransferase, N-acetyltransferaser og sulfotransferaser.
Opregulationen af HO-1, som er en virkelig beskyttende receptor med en potentiel vækst af CO, der i forbindelse med NO tillader vasodilation af iskæmiske celler.
Reduktion af jernoverbelastning gennem forhøjet ferritin og bilirubin som en lipofil antioxidant. Både fase II proteiner sammen med antioxidanterne er i stand til at løse det kroniske oxidative stress og også at genoplive et normalt redox system.
GSK3? under ledelse af AKT og PI3K, fosforylerer Fyn, hvilket resulterer i Fyn nuklear lokalisering, som Fyn fosforylerer Nrf2Y568, hvilket fører til nuklear eksport og nedbrydning af Nrf2.
NRF2 dæmper også TH1 / TH17 responsen og beriger TH2 responsen.
HDAC-hæmmere udløste Nrf2 signalvejen og opregulerede, at Nrf2 nedstrøms mål HO-1, NQO1 og glutamat-cystein-ligasekatalytisk underenhed eller GCLC ved at bremse Keap1 og opmuntre dissociation af Keap1 fra Nrf2, nrf2 nuklear translokation og Nrf2 -ARE binding.
Nrf2 indeholder en halveringstid på ca. 20 minutter under basale betingelser.
Mindsker IKK? pool gennem Keap1-binding reducerer I? B? nedbrydning og kan være den undvigende mekanisme, ved hvilken det er bevist, at Nrf2-aktivering inhiberer NF? B-aktivering.
Keap1 behøver ikke altid at være nedreguleret for at få NRF2 til at virke, såsom chlorophyllin, blåbær, ellaginsyre, astaxanthin og tepiphenoler kan øge NRF2 og KEAP1 ved 400 procent.
Nrf2 regulerer negativt gennem termen af stearoyl CoA desaturase eller SCD og citrat lyase eller CL.
Genetik
KEAP1
rs1048290
C allel - viste en betydelig risiko for og en beskyttende virkning mod resistent epilepsi (DRE)
rs11085735 (jeg er AC)
forbundet med nedgangshastighed i lungefunktionen i LHS
MAPT
rs242561
T-allel - beskyttende allel til Parkinson-sygdomme - havde stærkere NRF2 / sMAF-binding og var forbundet med de højere MAPT-mRNA-niveauer i 3-forskellige regioner i hjernen, herunder cerebellar cortex (CRBL), temporal cortex (TCTX), intralobulær hvidt stof (WHMT)
NFE2L2 (NRF2)
rs10183914 (jeg er CT)
T allel - Forøgede niveauer af Nrf2 protein og forsinket alder af Parkinsons begyndelse med fire år
rs16865105 (jeg er AC)
C allel - havde højere risiko for Parkinsons sygdom
rs1806649 (jeg er CT)
C allel - er blevet identificeret og kan være relevant for brystkræftets ætiologi.
forbundet med øget risiko for hospitalsindlæggelser i perioder med høje PM10 niveauer
rs1962142 (jeg er GG)
T allel - var forbundet med et lavt niveau af cytoplasmatisk NRF2-ekspression (P = 0.036) og negativ sulfuredoxinekspression (P = 0.042)
En allelbeskyttet nedgang i underarmsblodstrømmen (FEV) (tvungen ekspirationsvolumen på et sekund) i forhold til status for cigaretrygning (p = 0.004)
rs2001350 (jeg er TT)
T allel - beskyttet mod FEV-tilbagegang (tvunget ekspirationsvolumen på et sekund) i forhold til status for cigaretrygning (p = 0.004)
rs2364722 (jeg er AA)
En allelbeskyttet mod FEV-tilbagegang (tvunget ekspirationsvolumen på et sekund) i forhold til cigaretrygningsstatus (p = 0.004)
rs2364723
C allel - associeret med signifikant reduceret FEV hos japanske rygere med lungekræft
rs2706110
G allel - viste en betydelig risiko for og en beskyttende virkning mod resistent epilepsi (DRE)
AA alleler - viste signifikant reduceret KEAP1 ekspression
AA alleler - var forbundet med en øget risiko for brystkræft (P = 0.011)
rs2886161 (jeg er TT)
T allel - forbundet med Parkinsons sygdom
rs2886162
En allel – var forbundet med lav NRF2-ekspression (P = 0.011; OR, 1.988; CI, 1.162�3.400), og AA-genotypen var forbundet med en dårligere overlevelse (P = 0.032; HR, 1.687; CI, 1.047�)
rs35652124 (jeg er TT)
En allel - associeret med højere forbundet med alder ved starten for Parkinsons sygdom vs G allel
C allel - havde øget NRF2 protein
T allel - havde mindre NRF2-protein og større risiko for hjertesygdomme og blodtryk
rs6706649 (jeg er CC)
C allel - havde lavere NRF2 protein og øget risiko for Parkinsons sygdom
rs6721961 (jeg er GG)
T allel - havde lavere NRF2 protein
TT alleler - sammenhæng mellem cigaretrygning i tunge rygere og et fald i sædkvaliteten
TT-allel – var forbundet med øget risiko for brystkræft [P = 0.008; OR, 4.656; konfidensinterval (CI), 1.350–16.063] og T-allelen var forbundet med en lav grad af NRF2-proteinekspression (P = 0.0003; OR, 2.420; CI, 1.491–3.926) og negativ SRXN1-ekspression (P = 0.047; OR,1.867. 1.002; CI = 3.478�XNUMX)
T allel-allel blev også nominelt forbundet med ALI-relateret 28-dages mortalitet efter systemisk inflammatorisk respons syndrom
T allel - beskyttet mod FEV-tilbagegang (tvunget ekspirationsvolumen på et sekund) i forhold til status for cigaretrygning (p = 0.004)
G allel - forbundet med øget risiko for ALI efter større traumer hos europæiske og afrikanske amerikanere (odds ratio, OR 6.44; 95% konfidensinterval
AA alleler - forbundet med infektionsinduceret astma
AA alleler - udviste signifikant reduceret NRF2 genekspression og følgelig en øget risiko for lungekræft, især dem, der nogensinde havde røget
AA alleler - havde en signifikant højere risiko for at udvikle T2DM (ELLER 1.77; 95% CI 1.26, 2.49; p = 0.011) i forhold til dem med CC genotypen
AA alleler - stærk tilknytning mellem sårreparation og sen forgiftning af stråling (forbundet med en signifikant højere risiko for udvikling af forsinkede effekter hos afroamerikanere med en tendens i kaukasiere)
forbundet med oral østrogenbehandling og risiko for venøs tromboembolisme hos postmenopausale kvinder
rs6726395 (jeg er AG)
En allelbeskyttet mod FEV1-tilbagegang (tvunget ekspirationsvolumen på et sekund) i forhold til cigaretrygningsstatus (p = 0.004)
En allel - associeret med signifikant reduceret FEV1 hos japanske rygere med lungekræft
GG alleler - havde højere NRF2 niveauer og nedsat risiko for makuladegeneration
GG alleler - havde højere overlevelse med cholangiocarcinom
rs7557529 (jeg er CT)
C allel - forbundet med Parkinsons sygdom
Oxidativ stress og andre stressorer kan forårsage celleskader, som eventuelt kan føre til en række sundhedsmæssige problemer. Forskningsundersøgelser har vist, at Nrf2-aktivering kan fremme menneskekroppen's beskyttende antioxidantmekanisme, men forskere har diskuteret, at Nrf2 overekspression kan have enorme risici for overordnet sundhed og velvære. Forskellige former for kræft kan også forekomme med Nrf2-overaktivering.
Dr. Alex Jimenez DC, CCST Insight
Sulforaphane og dens virkninger på kræft, dødelighed, aldring, hjerne og adfærd, hjertesygdomme og mere
Isothiocyanater er nogle af de vigtigste planteforbindelser, du kan få i din kost. I denne video gør jeg det mest omfattende tilfælde til dem, der nogensinde er blevet lavet. Kort opmærksomhed span? Spring til dit yndlingsemne ved at klikke på et af nedenstående tidspunkter. Fuld tidslinje nedenfor.
Nøglesektioner:
00: 01: 14 - Kræft og dødelighed
00: 19: 04 - Aging
00: 26: 30 - Hjerne og adfærd
00: 38: 06 - Final recap
00: 40: 27 - Dosis
Fuld tidslinje:
00: 00: 34 - Introduktion af sulforaphane, et vigtigt fokus i videoen.
00: 01: 14 - Cruciferous vegetabilsk forbrug og reduktion af allårsag dødelighed.
00: 02: 12 - Prostatacancerrisiko.
00: 02: 23 - Kræftrisiko.
00: 02: 34 - Lungekræft i rygere risiko.
00: 02: 48 - Brystkræftrisiko.
00: 03: 13 - Hypotetisk: Hvad hvis du allerede har kræft? (Interventionel)
00: 03: 35 - Plausibel mekanisme, der driver kræft- og dødelighedsassosierende data.
00: 04: 38 - Sulforaphane og kræft.
00: 05: 32 - Dyrebevis, der viser stærk virkning af broccoli-spireekstrakt på udviklingen af blæretumor hos rotter.
00: 06: 06 - Virkning af direkte tilsætning af sulforaphan hos patienter med prostatacancer.
00: 07: 09 - Bioakkumulering af isothiocyanatmetabolitter i egentligt brystvæv.
00: 08: 32 - Inhibering af brystcancerstamceller.
00: 08: 53 - Historie lektion: brassicas blev etableret som at have sundhedsegenskaber selv i det gamle Rom.
00: 09: 16 - Sulforaphans evne til at øge kræftfremkaldende udskillelse (benzen, acrolein).
00: 09: 51 - NRF2 som en genetisk switch via antioxidant responselementer.
00: 10: 10 - Hvordan NRF2 aktivering øger kræftfremkaldende udskillelse via glutathion-S-konjugater.
00: 10: 34 - Spiraler øger glutathion-S-transferase og reducerer DNA-beskadigelse.
00: 11: 20 - Broccoli-spire drikker øger benzenudskillelsen med 61%.
00: 13: 31 - Broccoli spirehomogenat øger antioxidant enzymer i den øvre luftvej.
00: 15: 45 - Cruciferous vegetabilsk forbrug og dødelighed i hjertesygdomme.
00: 16: 55 - Broccoli spire pulver forbedrer blodlipider og overordnet hjertesygdomsrisiko hos type 2 diabetikere.
00: 19: 04 - Begyndelse af aldringssektionen.
00: 19: 21 - Sulforaphane beriget kost forbedrer levetiden for biller fra 15 til 30% (under visse forhold).
00: 20: 34 - Betydningen af lav inflammation for lang levetid.
00: 22: 05 - Cruciferous grøntsager og broccoli spire pulver synes at reducere et bredt udvalg af inflammatoriske markører hos mennesker.
00: 24: 14 - Musestudier tyder på, at sulforaphan kan forbedre adaptiv immunfunktion i alderdommen.
00: 25: 18 - Sulforaphane forbedrede hårvækst i en musmodel af skaldethed. Billede på 00: 26: 10.
00: 26: 30 - Begyndelse af hjerne- og adfærdssektionen.
00: 27: 18 - Effekt af broccoli-spireekstrakt på autisme.
00: 27: 48 - Virkning af glucoraphanin på skizofreni.
00: 28: 17 - Start af depression diskussion (sandsynlig mekanisme og undersøgelser).
00: 31: 21 - Mus undersøgelse ved hjælp af 10 forskellige modeller af stress-induceret depression viser sulforaphan på samme måde som fluoxetin (prozac).
00: 32: 00 - Undersøgelse viser direkte indtagelse af glucoraphanin hos mus er tilsvarende effektiv til forebyggelse af depression fra social nederlagsspændingsmodel.
00: 33: 01 - Begyndelsen af neurodegenerationsafsnittet.
00: 33: 30 - Sulforaphane og Alzheimers sygdom.
00: 33: 44 - Sulforaphane og Parkinsons sygdom.
00: 33: 51 - Sulforaphane og Hungtington's sygdom.
00: 35: 55 - Sulforaphane og neuronal plasticitet.
00: 36: 32 - Sulforaphane forbedrer læring som model af type II diabetes hos mus.
00: 37: 19 - Sulforaphane og duchenne muskeldystrofi.
00: 37: 44 - Myostatininhibering i muskel-satellitceller (in vitro).
00: 38: 06 - Late video recap: dødelighed og kræft, DNA-beskadigelse, oxidativ stress og betændelse, udslip af benzen, kardiovaskulær sygdom, type II diabetes, hjernens virkninger (depression, autisme, skizofreni, neurodegeneration), NRF2-pathway.
00: 40: 27 - Tanker om at finde ud af en dosis broccolispirer eller sulforaphane.
00: 41: 01 - Anecdoter på spiring hjemme.
00: 43: 14 - På tilberedningstemperaturer og sulforafanaktivitet.
00: 43: 45 - Gut-bakteriekonvertering af sulforaphan fra glucoraphanin.
00: 44: 24 - Tilskud fungerer bedre, når de kombineres med aktiv myrosinase fra grøntsager.
00: 44: 56 - Madlavningsteknikker og cruciferous grøntsager.
00: 46: 06 - Isothiocyanater som goitrogener.
Ifølge forskningsundersøgelser er Nrf2 en fundamental transkriptionsfaktor, som aktiverer cellernes beskyttende antioxidantmekanismer for at afgifte den menneskelige krop. Overekspressionen af Nrf2 kan dog forårsage sundhedsproblemer. Omfanget af vores information er begrænset til kiropraktik og rygsøjlesundhedsproblemer. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på�915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emnediskussion: Akut rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og ubesvarede arbejdsdage over hele verden. Rygsmerter tilskrives den næst hyppigste årsag til lægehjælp, der kun overtales af infektioner i øvre luftveje. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Rygsøjlen er en kompleks struktur, der består af knogler, led, ledbånd og muskler, blandt andet blødt væv. Skader og / eller forværrede forhold, såsom herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
Mange nuværende undersøgelser af kræft har gjort det muligt for sundhedspersonale at forstå den måde kroppen aftar. Ved at analysere opregulerede gener i tumorceller, opdagede forskerne nukleær erythroid 2-relateret faktor 2 signalveje, bedst kendt som Nrf2. NRF2 er en vigtig transkriptionsfaktor, som aktiverer menneskekroppen beskyttende antioxidantmekanismer for at regulere oxidation fra både eksterne og interne faktorer for at forhindre øgede niveauer af oxidativ stress.
Principper for Nrf2
NRF2 er afgørende for at opretholde den generelle sundhed og velvære, fordi det tjener det primære formål med at regulere, hvordan vi administrerer alt, hvad vi udsættes for dagligt og ikke bliver syge. NRF2-aktivering spiller en rolle i fase II-afgiftningssystemet. Fase II-afgiftning tager lipofile eller fedtopløselige frie radikaler og omdanner dem til hydrofile eller vandopløselige stoffer til udskillelse, mens de inaktiverer usædvanligt reaktive metabolitter og kemikalier som en konsekvens af fase I.
NRF2-aktivering reducerer den generelle oxidation og inflammation i den menneskelige krop gennem en hormonisk effekt. For at udløse NRF2 skal der opstå en inflammatorisk reaktion på grund af oxidation for at cellerne skal producere et adaptivt respons og skabe antioxidanter, såsom glutathion. For at nedbryde princippet om Nrf2 aktiverer oxidativt stress i det væsentlige NRF2, som derefter aktiverer et antioxidantrespons i den menneskelige krop. NRF2 funktioner til at afbalancere redox signalering eller ligevægten af oxidant og antioxidant niveauer i cellen.
En god illustration af, hvordan denne proces fungerer, kan demonstreres med træning. Gennem hver træning tilpasser muskelen sig, så den kan rumme endnu en træningssession. Hvis NRF2 bliver under- eller overudtrykt på grund af kroniske infektioner eller øget eksponering for toksiner, som kan observeres hos patienter, der har kronisk inflammatorisk respons syndrom eller CIRS, kan sundhedsmæssige problemer forværres efter NRF2-aktivering. Frem for alt, hvis DJ-1 bliver overoxideret, slutter NRF2-aktivering hurtigt for hurtigt.
Virkninger af NRF2-aktivering
NRF2 aktivering er stærkt udtrykt i lunger, lever og nyrer. Nukleær erythroid 2-relateret faktor 2 eller NRF2 fungerer mest ved at modvirke forøgede niveauer af oxidation i den menneskelige krop, som kan føre til oxidativ stress. Nrf2 aktivering kan hjælpe med at behandle en lang række sundhedsproblemer, men overaktivering af Nrf2 kan forværre forskellige problemer, som er demonstreret nedenfor.
Periodisk aktivering af Nrf2 kan hjælpe:
Aldring (dvs. lang levetid)
Autoimmunitet og samlet inflammation (dvs. arthritis, autisme)
Kræft og kemisk beskyttelse (dvs. EMF eksponering)
Depression og angst (dvs. PTSD)
Narkotikaeksponering (Alkohol, NSAID)
Udøvelse og udholdenhed præstationer
Gut Disease (dvs. SIBO, Dysbiose, Ulcerativ Colitis)
Hjertetransplantation (mens åbent NRF2 kan være dårligt, kan NRF2 hjælpe med reparation)
Hepatitis C
Nefritis (alvorlige tilfælde)
Vitiligo
Endvidere kan NRF2 hjælpe med at få specifikke kosttilskud, stoffer og medicin til at fungere. Mange naturlige tilskud kan også hjælpe med at udløse NRF2. Gennem aktuelle forskningsundersøgelser har forskere vist, at et stort antal forbindelser, der engang blev antaget at være antioxidanter, virkelig var oxidanter. Det er fordi næsten alle har brug for NRF2 for at fungere, selv kosttilskud som curcumin og fiskeolie. Kakao viste sig for eksempel at generere antioxidantvirkninger hos mus, der besidder NRF2-genet.
Måder at aktivere NRF2
I tilfælde af neurodegenerative sygdomme som Alzheimers sygdom, Parkinsons sygdom, slagtilfælde eller endda autoimmune sygdomme, er det nok bedst at have Nrf2 upreguleret, men på en hormonisk måde. Blanding af NRF2-aktivatorer kan også have en additiv eller synergistisk virkning, idet det lejlighedsvis kan være dosisafhængigt. De øverste måder at øge Nrf2 udtryk på er anført nedenfor:
HIST (motion) + CoQ10 + Sun (disse synergier meget godt)
Broccoli Spirer + LLLT på mit hoved og tarm
Butyrat + Super Kaffe + Morgen Sol
Akupunktur (dette er en alternativ metode, laser akupunktur kan også bruges)
Fastende
Cannabidiol (CBD)
Lions Mane + Melatonin
Alfa-lipoinsyre + DIM
Malurt
PPAR-gamma-aktivering
Følgende omfattende liste, der indeholder over 350 andre måder at aktivere Nrf2 på gennem diæt, livsstil og udstyr, probiotika, kosttilskud, urter og olier, hormoner og neurotransmittere, lægemidler/medicin og kemikalier, veje/transskriptionsfaktorer, samt andre måder, er kun en kort guide til, hvad der kan udløse Nrf2. For korthedens skyld i denne artikel har vi udeladt over 500 andre fødevarer, kosttilskud og forbindelser, som kan hjælpe med at aktivere Nrf2. Følgende er anført nedenfor:
Kost:
Acai bær
Alkohol (rødvin er bedre, især hvis der er en kork i det, da protocatechuicaldehyd fra korker også kan aktivere NRF2. Generelt anbefales ikke alkohol, selvom akut indtagelse øger NRF2. Kronisk indtagelse kan nedsætte NRF2.
Alger (kelp)
æbler
Sort te
Paranødder
Broccolispirer (og andre isothiocyanater, sulforaphane samt korsblomstrede grøntsager som bok choy, der har D3T)
Blåbær (0.6-10 g / dag)
Gulerødder (falcarinone)
Cayenne Pepper (Capsaicin)
Selleri (butylphthalid)
Chaga (Betulin)
Kamille te
Del
Kinesisk kartoffel
Chokeberries (Aronia)
Chokolade (mørk eller kakao)
Kanel
Kaffe (såsom chlorogensyre, Cafestol og Kahweol)
Cordyceps
Fisk (og skaldyr)
Hørfrø
Hvidløg
Ghee (muligvis)
Ingefær (og kardemonin)
Gojiberries
Grapefrugt (Naringenin - 50 mg / kg / d naringenin)
vindruer
Grøn te
Guava
Heart Of Palm
Hijiki / Wakame
Honeycomb
Kiwi
Bælgplanter
Lion's Mane
Mahuwa
Mangos (Mangiferin)
Mangosteen
Mælk (ged, ko-via regulering af mikrobiom)
morbær
Olivenolie (pomace - hydroxytyrosol og oleanolsyre)
Omega 6 fedtsyrer (Lipoxin A4)
Osange appelsiner (Morin)
Østers champignon
Papaya
Jordnødder
Duer Ærter
Granatæble (Punicalagin, Ellaginsyre)
Propolis (Pinocembrin)
Lilla søde kartofler
Rambutan (Geraniin)
Løg
Reishi
Rhodiola Rosea (Salidroside)
Risklid (cycloartenyl ferulat)
Riceberry
Rooibos Tea
Rosemary
Legend
Saflor
Sesame Oil
Soja (og isoflavoner, Daidzein, Genistein)
Squash
jordbær
Tartary boghvede
Timian
tomater
Tonka Bønner
Gurkemeje
Wasabi
Vandmelon
Livsstil og enheder:
Akupunktur og elektroakupunktur (via kollagenkaskade på ECM)
Blåt lys
Brainspil (øger NRF2 i hippocampus)
Caloric Restriction
Kold (brusere, dyk, isbad, udstyr, kryoterapi)
EMF'er (lav frekvens, såsom PEMF)
Motion (akut træning som HIST eller HIIT synes at være mere gavnlig for at inducere NRF2, mens længere træning ikke inducerer NRF2, men øger glutathionniveauerne)
Høj fedt kost (kost)
High Heat (Sauna)
Hydrogen Inhalation og Hydrogen Water
Hyperbaric Oxygen Therapy
Infrarød terapi (som Joovv)
Intravenøs C-vitamin
Ketogen diæt
Ozon
Rygning (anbefales ikke - akut rygning øger NRF2, kronisk rygning nedsætter NRF2. Hvis du vælger at ryge, kan Holy Basil hjælpe med at beskytte mod nedregulering af NRF2)
Sol (UVB og Infrarød)
Probiotika:
Bacillus subtilis (fmbJ)
Clostridiumbutyricum (MIYAIRI 588)
Lactobacillus brevis
Lactobacillus casei (SC4 og 114001)
Lactobacillus collinoides
Lactobacillus gasseri (OLL2809, L13-Ia og SBT2055)
Lactobacillus helveticus (NS8)
Lactobacillus paracasei (NTU 101)
Lactobacillus plantarum (C88, CAI6, FC225, SC4)
Lactobacillus rhamnosus (GG)
Kosttilskud, urter og olier:
Acetyl-L-carnitin (ALCAR) og carnitin
Allicin
Alpha-liposyre
amentoflavon
Andrographis paniculata
agmatin
apigenin
Arginin
Artiskok (Cyanropicrin)
Ashwaganda
Astragalus
Bacopa
Beefsteak (Isogemaketon)
Berberin
Beta-caryophyllen
Bidens Pilosa
Sort Cumin Seed Oil (Thymoquinon)
Boswellia
Butein
butyrat
Cannabidiol (CBD)
Carotenioider (som Beta-caroten [synergi med lycopen – 2 � 15 mg/d lycopen], Fucoxanthin, Zeaxanthin, Astaxanthin og Lutein)
Chitrak
Chlorella
klorofyl
Krysantemum zawadskii
Cinnamomea
Fælles Sundew
Kobber
Coptis
CoQ10
Curcumin
Damiana
Dan Shen / Red Sage (Miltirone)
DIM
dioscin
Dong Ling Cao
Dong Quai (kvindelig ginseng)
Ecklonia Cava
EGCG
Elecampane / Inula
Eucommia Bark
Ferulic Acid
fisetin
Fiskeolie (DHA / EPA - 3 1 g / d fiskeolie indeholdende 1098 mg EPA og 549 mg DHA)
Galangal
Gastrodin (Tian Ma)
Gentiana
Geranium
Ginkgo Biloba (Ginkgolide B)
kveller
Gotu Kola
Grape Seed Extract
Håret Agrimony
Haritaki (Triphala)
Hawthorn
Helichrysum
Henna (Juglone)
Hibiscus
Higenamine
Hellige Basil / Tulsi (Ursolsyre)
Humle
Horny Goat Weed (Icariin / Icariside)
Indigo Naturalis
Jern (anbefales ikke medmindre det er nødvendigt)
I3C
Jobs tårer
Moringa Oleifera (såsom Kaempferol)
Inchinkoto (combo of Zhi Zi og Wormwood)
Kudzu Root
Licorice Root
Lindera Root
Luteolin (høje doser til aktivering, lavere doser hæmmer NRF2 i kræft dog)
Magnolia
Manjistha
Maximowiczianum (Acerogenin A)
Mexicansk Arnica
Milk Thistle
MitoQ
Mu Xiang
Mucuna pruriens
Nicotinamid og NAD +
Panax Ginseng
Passionsblomst (såsom Chrysin, men chyrisin kan også reducere NRF2 via dysregulering af PI3K/Akt-signalering)
Resveratrol (Piceid og andre phytoøstrogener i det væsentlige, Knotweed)
Rose Hips
Palisander
Rutin
Sappanwood
Sarsaparilla
Saururus chinensis
SC-E1 (Gypsum, Jasmine, Licorice, Kudzu og Ballonblomst)
Schisandra
Selvhelbredelse (prunella)
Skullcap (Baicalin og Wogonin)
Får Sorrel
Si Wu Tang
Sideritis
Spikenard (Aralia)
Spirulina
St. John's Wort
sulforaphane
Sutherlandia
Tao Hong Si Wu
Taurin
Thunder God Vine (Triptolide)
Tocopheroler (såsom E-vitamin eller Linalool)
Tribulus R
Tu Si Zi
TUDCA
Vitamin A (selvom andre retinoider hæmmer NRF2)
C-vitamin (kun høj dosis, lav dosis hæmmer�NRF2)
Vitex / Chaste Tree
Hvid pæon (Paeoniflorin fra Paeonia lactiflora)
Malurt (Hispidulin og Artemisinin)
Xiao Yao Wan (Free and Easy Wanderer)
Yerba Santa (Eriodictyol)
Yuan Zhi (Tenuigin)
Zi Cao (vil reducere NRF2 i kræft)
Zink
Ziziphus Jujube
Hormoner og neurotransmittere:
adiponectin
Adropin
Østrogen (men kan nedsætte NRF2 i brystvæv)
Melatonin
Progesteron
Quinolinsyre (i beskyttende respons for at forhindre excitotoksicitet)
Serotonin
Skjoldbruskkirormoner som T3 (kan øge NRF2 i raske celler, men mindske det i kræft)
Vitamin D
Narkotika / medicin og kemikalier:
Acetaminophen
Acetazolamid
Amlodipin
auranofin
Bardoxolonmethyl (BARD)
benznidazol
BHA
CDDO-imidazolid
Ceftriaxon (og beta-lactam antibiotika)
Cialis
Dexamethason
Diprivan (Propofol)
eriodictyol
Exendin-4
ezetimibe
Fluorid
fumarat
HNE (oxideret)
idazoxan
Uorganisk arsen og natrium arsenit
JQ1 (kan også hæmme NRF2, ukendt)
Letairis
melphalan
Methazolamid
Methylenblå
Nifedipin
NSAID
oltipraz
PPI'er (såsom omeprazol og lansoprazol)
Protandim - gode resultater in vivo, men svage / ikke-eksisterende ved aktivering af NRF2 hos mennesker
probucol
Rapamycin
reserpin
ruthenium
sitaxentan
Statiner (såsom Lipitor og Simvastatin)
Tamoxifen
Tang Luo Ning
tBHQ
Tecfidera (dimethylfumarat)
THC (ikke så stærk som CBD)
Theophyllin
umbelliferon
Ursodeoxycholsyre (UDCA)
Verapamil
Viagra
4-Acetoxyphenol
Stier / Transkriptionsfaktorer:
?7 nAChR-aktivering
AMPK
Bilirubin
CDK20
CKIP-1
CYP2E1
EAAT'er
Gankyrin
Gremlin
GJA1
H-ferritin ferroxidase
HDAC-hæmmere (såsom valproinsyre og TSA, men kan forårsage NRF2-ustabilitet)
Heat Shock Proteins
IL-17
IL-22
Klotho
lad-7 (slår ned mBach1 RNA)
MAPK
Michael acceptorer (mest)
miR-141
miR-153
miR-155 (slår også mBach1 RNA)
miR-7 (i hjernen, hjælper mod kræft og skizofreni)
Notch1
Oxidativ stress (såsom ROS, RNS, H2O2) og elektrofiler
PGC-1?
PKC-delta
PPAR-gamma (synergistiske effekter)
Sigma-1-receptorinhibering
SIRT1 (øger NRF2 i hjernen og lungerne, men kan reducere det generelt)
SIRT2
SIRT6 (i lever og hjerne)
SRXN1
TrxR1-hæmning (såvel dæmpning eller udtømning)
Zink protoporphyrin
4-HHE
Andre:
Ankaflavin
Asbest
Aviciner
Bacillus amyloliquefaciens (bruges i landbruget)
Kulilte
Daphnetin
Glutathiondepletering (udtømning på 80%–90% muligvis)
Gymnaster koraiensis
Hepatitis C
Herpes (HSV)
Indisk asketræ
Indigowood rod
Isosalipurposide
Isorhamentin
Monascin
Omaveloxolone (stærk, aka RTA-408)
PDTC
Selenmangel (selenmangel kan øge NRF2)
Sibirisk lærk
Sophoraflavanon G
Tadehagi triquetrum
Toona sinensis (7-DGD)
Trompetblomst
63171 og 63179 (stærk)
Den nukleare erythroid 2-relaterede faktor 2 signalvej, der er bedst kendt af akronym Nrf2, er en transkriptionsfaktor, som spiller den vigtigste rolle at regulere de beskyttende antioxidantmekanismer i menneskekroppen, især for at kontrollere oxidativ stress. Mens øgede niveauer af oxidativ stress kan aktivere Nrf2, forbedres dets virkninger kraftigt gennem tilstedeværelsen af specifikke forbindelser. Visse fødevarer og kosttilskud hjælper med at aktivere Nrf2 i menneskekroppen, herunder isothiocyanat sulforaphan fra broccoli spirer. Dr. Alex Jimenez DC, CCST Insight
Sulforaphane og dens virkninger på kræft, dødelighed, aldring, hjerne og adfærd, hjertesygdomme og mere
Isothiocyanater er nogle af de vigtigste planteforbindelser, du kan få i din kost. I denne video gør jeg det mest omfattende tilfælde til dem, der nogensinde er blevet lavet. Kort opmærksomhed span? Spring til dit yndlingsemne ved at klikke på et af nedenstående tidspunkter. Fuld tidslinje nedenfor.
Nøglesektioner:
00: 01: 14 - Kræft og dødelighed
00: 19: 04 - Aging
00: 26: 30 - Hjerne og adfærd
00: 38: 06 - Final recap
00: 40: 27 - Dosis
Fuld tidslinje:
00: 00: 34 - Introduktion af sulforaphane, et vigtigt fokus i videoen.
00: 01: 14 - Cruciferous vegetabilsk forbrug og reduktion af allårsag dødelighed.
00: 02: 12 - Prostatacancerrisiko.
00: 02: 23 - Kræftrisiko.
00: 02: 34 - Lungekræft i rygere risiko.
00: 02: 48 - Brystkræftrisiko.
00: 03: 13 - Hypotetisk: Hvad hvis du allerede har kræft? (Interventionel)
00: 03: 35 - Plausibel mekanisme, der driver kræft- og dødelighedsassosierende data.
00: 04: 38 - Sulforaphane og kræft.
00: 05: 32 - Dyrebevis, der viser stærk virkning af broccoli-spireekstrakt på udviklingen af blæretumor hos rotter.
00: 06: 06 - Virkning af direkte tilsætning af sulforaphan hos patienter med prostatacancer.
00: 07: 09 - Bioakkumulering af isothiocyanatmetabolitter i egentligt brystvæv.
00: 08: 32 - Inhibering af brystcancerstamceller.
00: 08: 53 - Historie lektion: brassicas blev etableret som at have sundhedsegenskaber selv i det gamle Rom.
00: 09: 16 - Sulforaphans evne til at øge kræftfremkaldende udskillelse (benzen, acrolein).
00: 09: 51 - NRF2 som en genetisk switch via antioxidant responselementer.
00: 10: 10 - Hvordan NRF2 aktivering øger kræftfremkaldende udskillelse via glutathion-S-konjugater.
00: 10: 34 - Spiraler øger glutathion-S-transferase og reducerer DNA-beskadigelse.
00: 11: 20 - Broccoli-spire drikker øger benzenudskillelsen med 61%.
00: 13: 31 - Broccoli spirehomogenat øger antioxidant enzymer i den øvre luftvej.
00: 15: 45 - Cruciferous vegetabilsk forbrug og dødelighed i hjertesygdomme.
00: 16: 55 - Broccoli spire pulver forbedrer blodlipider og overordnet hjertesygdomsrisiko hos type 2 diabetikere.
00: 19: 04 - Begyndelse af aldringssektionen.
00: 19: 21 - Sulforaphane beriget kost forbedrer levetiden for biller fra 15 til 30% (under visse forhold).
00: 20: 34 - Betydningen af lav inflammation for lang levetid.
00: 22: 05 - Cruciferous grøntsager og broccoli spire pulver synes at reducere et bredt udvalg af inflammatoriske markører hos mennesker.
00: 24: 14 - Musestudier tyder på, at sulforaphan kan forbedre adaptiv immunfunktion i alderdommen.
00: 25: 18 - Sulforaphane forbedrede hårvækst i en musmodel af skaldethed. Billede på 00: 26: 10.
00: 26: 30 - Begyndelse af hjerne- og adfærdssektionen.
00: 27: 18 - Effekt af broccoli-spireekstrakt på autisme.
00: 27: 48 - Virkning af glucoraphanin på skizofreni.
00: 28: 17 - Start af depression diskussion (sandsynlig mekanisme og undersøgelser).
00: 31: 21 - Mus undersøgelse ved hjælp af 10 forskellige modeller af stress-induceret depression viser sulforaphan på samme måde som fluoxetin (prozac).
00: 32: 00 - Undersøgelse viser direkte indtagelse af glucoraphanin hos mus er tilsvarende effektiv til forebyggelse af depression fra social nederlagsspændingsmodel.
00: 33: 01 - Begyndelsen af neurodegenerationsafsnittet.
00: 33: 30 - Sulforaphane og Alzheimers sygdom.
00: 33: 44 - Sulforaphane og Parkinsons sygdom.
00: 33: 51 - Sulforaphane og Hungtington's sygdom.
00: 35: 55 - Sulforaphane og neuronal plasticitet.
00: 36: 32 - Sulforaphane forbedrer læring som model af type II diabetes hos mus.
00: 37: 19 - Sulforaphane og duchenne muskeldystrofi.
00: 37: 44 - Myostatininhibering i muskel-satellitceller (in vitro).
00: 38: 06 - Late video recap: dødelighed og kræft, DNA-beskadigelse, oxidativ stress og betændelse, udslip af benzen, kardiovaskulær sygdom, type II diabetes, hjernens virkninger (depression, autisme, skizofreni, neurodegeneration), NRF2-pathway.
00: 40: 27 - Tanker om at finde ud af en dosis broccolispirer eller sulforaphane.
00: 41: 01 - Anecdoter på spiring hjemme.
00: 43: 14 - På tilberedningstemperaturer og sulforafanaktivitet.
00: 43: 45 - Gut-bakteriekonvertering af sulforaphan fra glucoraphanin.
00: 44: 24 - Tilskud fungerer bedre, når de kombineres med aktiv myrosinase fra grøntsager.
00: 44: 56 - Madlavningsteknikker og cruciferous grøntsager.
00: 46: 06 - Isothiocyanater som goitrogener.
Ifølge mange aktuelle forskningsundersøgelser er den nukleare erythroid 2-relaterede faktor 2-signalvej, bedst kendt som Nrf2, en grundlæggende transkriptionsfaktor, der aktiverer cellernes beskyttende antioxidantmekanismer til at afgifte menneskekroppen fra både eksterne og interne faktorer og forhindrer øget niveauer af oxidativ stress. Omfanget af vores information er begrænset til kiropraktik og spinal sundhedsspørgsmål. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på 915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emnediskussion: Akut rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og ubesvarede arbejdsdage over hele verden. Rygsmerter tilskrives den næst hyppigste årsag til lægehjælp, der kun overtales af infektioner i øvre luftveje. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Rygsøjlen er en kompleks struktur, der består af knogler, led, ledbånd og muskler, blandt andet blødt væv. Skader og / eller forværrede forhold, såsom herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller bilulykkesskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe med at lindre rygsmerter gennem brug af spinaljusteringer og manuelle manipulationer, hvilket i sidste ende forbedrer smertelindring. �
Oxidativ stress er en væsentlig bidragyder til udviklingen af en række sundhedsproblemer, herunder kræft, hjertesygdomme, diabetes, accelereret aldring og neurodegeneration. Antioxidantrige fødevarer, urter og kosttilskud kan bruges til at beskytte den menneskelige krop mod høje niveauer af oxidativt stress. Nyere forskningsundersøgelser har vist, at Nrf2-genvej kan hjælpe med at forstærke virkningen af antioxidanter. Det fordelene ved Nrf2 er beskrevet nedenfor.
Beskytter kroppen mod toksiner
NRF2 er et iboende stof, som kan beskytte cellerne mod skadelige, interne og eksterne forbindelser. NRF2 kan hjælpe med at berige den menneskelige krops reaktion på lægemidler/medicin og toksiner og forbedre produktionen af proteiner, der hjælper med at eliminere forbindelser fra cellen, kendt som multidrug resistens-associerede proteiner eller MRP'er. NRF2 udløses for eksempel ved indånding af cigaretrøg for at give lungerne mulighed for at afgifte.
Derudover er det vigtigt for lungerne at beskytte sig mod allergener, virussygdomme, bakterielle endotoksiner, hyperoksi og forskellige miljøforurenende stoffer. Den konstante udløser af Nrf2 kan dog reducere niveauerne af et stof kendt som glutathion i hele menneskekroppen. NRF2 kan også beskytte leveren mod toksicitet, og det kan beskytte leveren mod arsen hepatotoksicitet. Desuden beskytter NRF2 leveren og hjernen mod alkoholforbrug. For eksempel kan Nrf2 beskytte mod acetaminophen-toksicitet.
Bekæmper betændelse og oxidativ stress
NRF2-aktivering kan hjælpe med at bekæmpe inflammation ved at mindske inflammatoriske cytokiner, såsom dem, der er til stede i psoriasis. NRF2 kan også mindske inflammation forbundet med en række sundhedsproblemer som gigt og fibrose i leveren, nyrerne og lungerne. NRF2 kan også hjælpe med at kontrollere allergier ved at sænke Th1/Th17 cytokiner og hæve TH2 cytokiner. Dette kan være gavnligt for lidelser som astma.
NRF2 beskytter desuden mod cellulær skade fra blåt lys�og mod UVA/UVB� fundet i sollys. Nrf2-mangler kan gøre det meget nemmere at blive solskoldet. En begrundelse bag dette er, at NRF2 er i stand til at regulere kollagen som reaktion på UV-stråling. Advanced Glycation End-Products, eller AGE'er, bidrager til udviklingen af mange sundhedsproblemer, herunder diabetes og neurodegenerative sygdomme. NRF2 kan mindske det oxidative stress af AGE'er i kroppen. NRF2 kan også beskytte den menneskelige krop mod højere niveauer af varmebaseret stress.
Forbedrer mitokondrier og træningsydelse
NRF2 er en mitokondriel booster. NRF2-aktivering bidrager til en stigning i ATP-energi for mitokondrier, foruden øget brug af ilt eller citrat og fedt. Uden NRF2 ville mitokondrier bare have evnen til at fungere med sukker eller glukose i stedet for fedt. NRF2 er også afgørende for, at mitokondrier kan udvikle sig gennem en proces kendt som biogenese. NRF2-aktivering er afgørende for at �drage fordel af� fordelene ved træning.
På grund af Nrf2's aktivitet øger træning mitokondriefunktionen, hvor dette resultat kan forstærkes med CoQ10, Cordyceps og kaloriebegrænsning. Moderat træning eller akut træning inducerer mitokondriel biogenese og en forhøjet syntese af superoxiddismutase eller SOD og hæm-oxygenase-1 eller HO-1 gennem NRF2-aktivering. Alpha-Lipoic Acid, eller ALA, og Dan Shen kan booste NRF2-medieret mitokondriel biogenese. Ydermere kan NRF2 også forbedre træningstolerancen, hvor NRF2-sletning gør træning skadeligt.
Beskytter mod hypoxi
NRF2 hjælper også med at beskytte den menneskelige krop mod cellulært ilttab/udtømning, et sundhedsproblem kaldet hypoxi. Personer med CIRS har reducerede niveauer af ilt, da deres NRF2 er blokeret, hvilket resulterer i reducerede niveauer af både VEGF, HIF1 og HO-1. Normalt overudtrykkes miR-101, som er nødvendig for dannelsen af stamceller, hos raske individer med hypoxi og øger mængden af NRF2/HO-1 og VEGF/eNOS, hvilket forhindrer hjerneskade, men det ser ikke ud til at forekomme. i CIRS.
Hypoxi, karakteriseret ved lav HIF1, i CIRS kan også resultere i en utæt blodhjernebarriere på grund af en NRF2-ubalance. Salidroside, der ligger i Rhodiola, fungerer på NRF2-aktivering og hjælper med hypoxi ved at øge niveauerne af VEGF og HIF1 i den menneskelige krop. NRF2 kan også i sidste ende beskytte mod ophobning af laktat i hjertet. NRF2-aktivering kan også stoppe hypoxi-induceret højdekørselssyge eller AMS.
Bremser aldring
Adskillige forbindelser, som kan være dødelige i store mængder, kan øge levetiden i ret små mængder på grund af xenohormesis gennem NRF2, PPAR-gamma og FOXO. En meget lille mængde toksiner øger cellens evne til at blive bedre rustet til næste gang den udfordres med et toksin, men dette er ikke en godkendelse til at indtage giftige kemikalier.
En god illustration af denne proces er med kaloriebegrænsning. NRF2 kan forbedre cellernes levetid ved at hæve deres niveauer af mitokondrier og antioxidanter samt sænke cellernes evne til at dø. NRF2 aftager med aldring, fordi NRF2 forhindrer stamceller i at dø og hjælper dem med at regenerere. NRF2 spiller en rolle i at forbedre sårheling.
Booster det vaskulære system
Udført korrekt med produktionen af sulforaphane, kan NRF2-aktivering beskytte mod hjertesygdomme som forhøjet blodtryk eller hypertension og åreforkalkning eller åreforkalkning. NRF2 kan øge Acetylcholins eller ACh, afslappende aktivitet på det vaskulære system, samtidig med at det reducerer kolesterol-induceret stress. Nrf2-aktivering kan styrke hjertet, men overaktiveret Nrf2 kan øge sandsynligheden for hjerte-kar-sygdomme.
Statiner kan forebygge eller føre til hjerte-kar-sygdomme. NRF2 spiller også en vigtig rolle i at balancere jern og calcium, som kan beskytte den menneskelige krop mod at have forhøjede niveauer af jern. For eksempel kan Sirtuin 2 eller SIRT2 regulere jernhomeostase i celler ved aktivering af NRF2, som menes at være påkrævet for sunde niveauer af jern. NRF2 kan også hjælpe med seglcellesygdom eller SCD. NRF2-dysfunktion kan være en årsag bag endotoksæmi som med dysbiose eller lectin-induceret hypertension. Nrf2 kan også beskytte den menneskelige krop mod amfetamin-induceret skade på det vaskulære system.
Bekæmper neuroinflammation
NRF2 kan skærme mod og hjælpe med betændelse i hjernen, almindeligvis omtalt som neuroinflammation. Ydermere kan NRF2 hjælpe med et udvalg af sygdomme i centralnervesystemet eller CNS, herunder:
Alzheimers sygdom (AD) – reducerer amyloid beta-stress på mitokondrier
Amyotrofisk lateral sklerose (ALS)
Huntingtons sygdom (HD)
Multipel sklerose (MS)
Nerveregenerering
Parkinsons sygdom (PD) – beskytter dopamin
Rygmarvsskade (SCI)
Slagtilfælde (iskæmisk og hæmoragisk) - hjælper med hypoxi
Traumatisk Hjerneskade
NRF2 har afsløret et fald i neuroinflammation hos teenagere med autismespektrumforstyrrelser eller ASD. Idebenone parrer korrekt med NRF2-aktivatorer i modsætning til neuroinflammation. NRF2 kan også forbedre Blood Brain Barrier,�eller BBB. For eksempel kan NRF2-aktivering med carnosinsyre opnået fra rosmarin og salvie krydse BBB og forårsage neurogenese. NRF2 har også vist sig at hæve Brain Derived Neurotrophic Factor eller BDNF.
NRF2 modulerer også nogle kosttilskuds evne til at forårsage nervevækstfaktor eller NGF, da det også kan hjælpe med hjernetåge og glutamat-inducerede problemer ved at modulere N-Methyl-D-Aspartat- eller NMDA-receptorer. Det kan også sænke det oxidative stress fra quinolinsyre, kaldet QUIN. NRF2-aktivering kan beskytte mod anfald, og store doser kan mindske randen af et anfald. Ved regelmæssige doser af stimulering kan NRF2 forbedre kognitive evner efter et anfald ved at sænke ekstracellulært glutamat i hjernen og ved dets evne til at trække cystein fra glutamat og glutathion.
Lindrer depression
Ved depression er det normalt at bemærke betændelse i hjernen, især fra den præfrontale cortex og hippocampus, samt nedsat BDNF. I nogle versioner af depression kan NRF2 forbedre depressive symptomer ved at sænke inflammation i hjernen og øge BDNF-niveauer. Agmatins evne til at mindske depression ved at hæve noradrenalin, dopamin, serotonin og BDNF i hippocampus afhænger af NRF2-aktivering.
Indeholder anti-cancer egenskaber
NRF2 er ligeledes en tumorundertrykker, da den er en tumorpromoter, hvis den ikke administreres i overensstemmelse hermed. NRF2 kan beskytte mod kræft forårsaget af frie radikaler og oxidativt stress, men NRF2-overekspression kan også findes i kræftceller. Intens aktivering af NRF2 kan hjælpe med en række kræftformer. For eksempel kan tilskuddet Protandim reducere hudkræft ved NRF2-aktivering.
Lindrer smerte
Gulf War Illness, eller GWI, en bemærkelsesværdig sygdom, der påvirker Golfkrigsveteraner, er en samling uforklarlige, kroniske symptomer, som kan omfatte træthed, hovedpine, ledsmerter, fordøjelsesbesvær, søvnløshed, svimmelhed, luftvejslidelser og hukommelsesproblemer. NRF2 kan forbedre symptomer på GWI ved at mindske hippocampus og generel inflammation, udover at mindske smerte. NRF2 kan desuden hjælpe med smerter fra kropslig nerveskade og forbedre nerveskader fra diabetisk neuropati.
Forbedrer diabetes
Høje glukoseniveauer, bedst omtalt som hyperglykæmi, forårsager oxidativ skade på cellerne på grund af forstyrrelse af mitokondriefunktionen. NRF2-aktivering kan beskytte den menneskelige krop mod hyperglykæmi's skade på cellen og derved forhindre celledød. NRF2-aktivering kan desuden beskytte, genoprette og forbedre bugspytkirtlens beta-celle funktion, samtidig med at insulinresistens reduceres.
Beskytter syn og hørelse
NRF2 kan beskytte mod skader på øjet fra diabetisk retinopati. Det kan også undgå dannelsen af grå stær og beskytte fotoreceptorer i modsætning til lysinduceret død. NRF2 beskytter desuden øret, eller cochlea, mod stress og høretab.
Kan hjælpe fedme
NRF2 kan hjælpe med fedme primært på grund af dets evne til at regulere variabler, der virker på fedtophobning i den menneskelige krop. NRF2-aktivering med sulforaphane kan hæve hæmningen af fedtsyresyntese eller FAS og frakoblingsproteiner eller UCP, hvilket resulterer i mindre fedtophobning og mere brunt fedt, karakteriseret som fedt, der omfatter flere mitokondrier.
Beskytter tarmen
NRF2 hjælper med at beskytte tarmen ved at beskytte tarmmikrobiomets homeostase. For eksempel vil lactobacillus probiotika udløse NRF2 for at beskytte tarmen mod oxidativt stress. NRF2 kan også hjælpe med at forhindre colitis ulcerosa eller UC.
Beskytter kønsorganer
NRF2 kan beskytte testiklerne og holde sædceller fra skade hos personer med diabetes. Det kan også hjælpe med erektil dysfunktion eller ED. Nogle libido-forstærkende kosttilskud som Mucuna, Tribulus og Ashwaganda kan forbedre den seksuelle funktion via NRF2-aktivering. Andre faktorer, der booster NRF2, såsom sollys eller broccolispirer, kan også hjælpe med at forbedre libido.
Regulerer knogler og muskler
Oxidativ stress kan resultere i knogletæthed og styrkereduktion, hvilket er normalt ved osteoporose. NRF2-aktivering kunne have evnen til at forbedre antioxidanter i knogler og beskytte mod ældning af knogler. NRF2 kan også forhindre muskeltab og forbedre Duchennes muskeldystrofi eller DMD.
Indeholder anti-virale egenskaber
Sidst, men ikke mindst, kan NRF2-aktivering i sidste ende hjælpe med at forsvare den menneskelige krop mod adskillige vira. Hos patienter med dengue-virus var symptomerne ikke så intense hos personer, der havde højere niveauer af NRF2 sammenlignet med personer, der havde mindre grader af NRF2. NRF2 kan også hjælpe mennesker, der har human immundefekt-1-virus eller HIV. NRF2 kan beskytte mod oxidativ stress fra Adeno-Associated Virus, eller AAV og H. Pylori. Endelig kan Lindera Root undertrykke hepatitis C-virus med NRF2-aktivering.
Nrf2, eller NF-E2-relateret faktor 2, er en transkriptionsfaktor fundet hos mennesker, som regulerer ekspressionen af et specifikt sæt antioxidant- og afgiftende gener. Denne signalvej aktiveres på grund af oxidativ stress, da den forbedrer talrige antioxidanter og fase II-leverafgiftningsenzymer for at genoprette homeostase i den menneskelige krop. Mennesker er tilpasset til at fungere gennem en tilstand af homeostase eller balance. Når kroppen konfronteres med oxidativt stress, aktiveres Nrf2 for at regulere oxidation og kontrollere den stress, den forårsager. Nrf2 er afgørende for at forhindre sundhedsproblemer forbundet med oxidativt stress. Dr. Alex Jimenez DC, CCST Insight
Sulforaphane og dens virkninger på kræft, dødelighed, aldring, hjerne og adfærd, hjertesygdomme og mere
Isothiocyanater er nogle af de vigtigste planteforbindelser, du kan få i din kost. I denne video gør jeg det mest omfattende tilfælde til dem, der nogensinde er blevet lavet. Kort opmærksomhed span? Spring til dit yndlingsemne ved at klikke på et af nedenstående tidspunkter. Fuld tidslinje nedenfor.
Nøglesektioner:
00: 01: 14 - Kræft og dødelighed
00: 19: 04 - Aging
00: 26: 30 - Hjerne og adfærd
00: 38: 06 - Final recap
00: 40: 27 - Dosis
Fuld tidslinje:
00: 00: 34 - Introduktion af sulforaphane, et vigtigt fokus i videoen.
00: 01: 14 - Cruciferous vegetabilsk forbrug og reduktion af allårsag dødelighed.
00: 02: 12 - Prostatacancerrisiko.
00: 02: 23 - Kræftrisiko.
00: 02: 34 - Lungekræft i rygere risiko.
00: 02: 48 - Brystkræftrisiko.
00: 03: 13 - Hypotetisk: Hvad hvis du allerede har kræft? (Interventionel)
00: 03: 35 - Plausibel mekanisme, der driver kræft- og dødelighedsassosierende data.
00: 04: 38 - Sulforaphane og kræft.
00: 05: 32 - Dyrebevis, der viser stærk virkning af broccoli-spireekstrakt på udviklingen af blæretumor hos rotter.
00: 06: 06 - Virkning af direkte tilsætning af sulforaphan hos patienter med prostatacancer.
00: 07: 09 - Bioakkumulering af isothiocyanatmetabolitter i egentligt brystvæv.
00: 08: 32 - Inhibering af brystcancerstamceller.
00: 08: 53 - Historie lektion: brassicas blev etableret som at have sundhedsegenskaber selv i det gamle Rom.
00: 09: 16 - Sulforaphans evne til at øge kræftfremkaldende udskillelse (benzen, acrolein).
00: 09: 51 - NRF2 som en genetisk switch via antioxidant responselementer.
00: 10: 10 - Hvordan NRF2 aktivering øger kræftfremkaldende udskillelse via glutathion-S-konjugater.
00: 10: 34 - Spiraler øger glutathion-S-transferase og reducerer DNA-beskadigelse.
00: 11: 20 - Broccoli-spire drikker øger benzenudskillelsen med 61%.
00: 13: 31 - Broccoli spirehomogenat øger antioxidant enzymer i den øvre luftvej.
00: 15: 45 - Cruciferous vegetabilsk forbrug og dødelighed i hjertesygdomme.
00: 16: 55 - Broccoli spire pulver forbedrer blodlipider og overordnet hjertesygdomsrisiko hos type 2 diabetikere.
00: 19: 04 - Begyndelse af aldringssektionen.
00: 19: 21 - Sulforaphane beriget kost forbedrer levetiden for biller fra 15 til 30% (under visse forhold).
00: 20: 34 - Betydningen af lav inflammation for lang levetid.
00: 22: 05 - Cruciferous grøntsager og broccoli spire pulver synes at reducere et bredt udvalg af inflammatoriske markører hos mennesker.
00: 24: 14 - Musestudier tyder på, at sulforaphan kan forbedre adaptiv immunfunktion i alderdommen.
00: 25: 18 - Sulforaphane forbedrede hårvækst i en musmodel af skaldethed. Billede på 00: 26: 10.
00: 26: 30 - Begyndelse af hjerne- og adfærdssektionen.
00: 27: 18 - Effekt af broccoli-spireekstrakt på autisme.
00: 27: 48 - Virkning af glucoraphanin på skizofreni.
00: 28: 17 - Start af depression diskussion (sandsynlig mekanisme og undersøgelser).
00: 31: 21 - Mus undersøgelse ved hjælp af 10 forskellige modeller af stress-induceret depression viser sulforaphan på samme måde som fluoxetin (prozac).
00: 32: 00 - Undersøgelse viser direkte indtagelse af glucoraphanin hos mus er tilsvarende effektiv til forebyggelse af depression fra social nederlagsspændingsmodel.
00: 33: 01 - Begyndelsen af neurodegenerationsafsnittet.
00: 33: 30 - Sulforaphane og Alzheimers sygdom.
00: 33: 44 - Sulforaphane og Parkinsons sygdom.
00: 33: 51 - Sulforaphane og Hungtington's sygdom.
00: 35: 55 - Sulforaphane og neuronal plasticitet.
00: 36: 32 - Sulforaphane forbedrer læring som model af type II diabetes hos mus.
00: 37: 19 - Sulforaphane og duchenne muskeldystrofi.
00: 37: 44 - Myostatininhibering i muskel-satellitceller (in vitro).
00: 38: 06 - Late video recap: dødelighed og kræft, DNA-beskadigelse, oxidativ stress og betændelse, udslip af benzen, kardiovaskulær sygdom, type II diabetes, hjernens virkninger (depression, autisme, skizofreni, neurodegeneration), NRF2-pathway.
00: 40: 27 - Tanker om at finde ud af en dosis broccolispirer eller sulforaphane.
00: 41: 01 - Anecdoter på spiring hjemme.
00: 43: 14 - På tilberedningstemperaturer og sulforafanaktivitet.
00: 43: 45 - Gut-bakteriekonvertering af sulforaphan fra glucoraphanin.
00: 44: 24 - Tilskud fungerer bedre, når de kombineres med aktiv myrosinase fra grøntsager.
00: 44: 56 - Madlavningsteknikker og cruciferous grøntsager.
00: 46: 06 - Isothiocyanater som goitrogener.
Når den menneskelige krop konfronteres med skadelige indre og ydre faktorer som toksiner, skal cellerne hurtigt udløse deres antioxidantevner for at modvirke oxidativt stress. Fordi øgede niveauer af oxidativt stress er blevet bestemt til at forårsage en række sundhedsproblemer, er det vigtigt at bruge Nrf2-aktivering for at drage fordel af dets fordele. Omfanget af vores information er begrænset til kiropraktik og rygsøjlesundhedsproblemer. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på�915-850-0900 .
Kurateret af Dr. Alex Jimenez
Yderligere emnediskussion: Akut rygsmerter
Rygsmerte er en af de mest udbredte årsager til handicap og ubesvarede arbejdsdage over hele verden. Rygsmerter tilskrives den næst hyppigste årsag til lægekontorbesøg, der kun er større end infektioner i de øvre luftveje. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Rygsøjlen er en kompleks struktur, der består af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette kan kvæstelser og / eller forværrede forhold, såsom herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller bilulykkesskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe med at lindre rygsmerter gennem brug af spinaljusteringer og manuelle manipulationer, hvilket i sidste ende forbedrer smertelindring. �
sulforaphane er et fytokemikalie, et stof inden for isothiocyanatgruppen af organiske svovlforbindelser, som findes i korsblomstrede grøntsager, såsom broccoli, kål, blomkål og rosenkål. Den kan også findes i bok choy, grønkål, collards, sennepsgrønt og brøndkarse. Forskningsundersøgelser har vist, at sulforaphane kan hjælpe med at forebygge forskellige former for kræft ved aktivering af produktionen af Nrf2, eller nuklear faktor erythroid 2-relateret faktor, en transkriptionsfaktor, som regulerer beskyttende antioxidantmekanismer, der kontrollerer cellens respons på oxidanter. Formålet med den følgende artikel er at beskrive sulforafans funktion.
Abstrakt
KEAP1-Nrf2-ARE antioxidantsystemet er et hovedmiddel, hvormed celler reagerer på oxidative og xenobiotisk stress. Sulforaphane (SFN), et elektrofilt isothiocyanat afledt af korsblomstrede grøntsager, aktiverer KEAP1-Nrf2-ARE-vejen og er blevet et molekyle af interesse i behandlingen af sygdomme, hvor kronisk oxidativt stress spiller en stor ætiologisk rolle. Vi demonstrerer her, at mitokondrierne af dyrkede, humane retinale pigmentepitelceller (RPE-1) behandlet med SFN gennemgår hyperfusion, der er uafhængig af både Nrf2 og dens cytoplasmatiske inhibitor KEAP1. Mitokondriel fusion er blevet rapporteret at være cytobeskyttende ved at hæmme poredannelse i mitokondrier under apoptose, og i overensstemmelse med dette viser vi Nrf2-uafhængig, cytobeskyttelse af SFN-behandlede celler udsat for apoptose-induceren, staurosporin. Mekanistisk mindsker SFN rekrutteringen og/eller tilbageholdelsen af den opløselige fissionsfaktor Drp1 til mitokondrier og til peroxisomer, men påvirker ikke den samlede Drp1-overflod. Disse data viser, at de gavnlige egenskaber ved SFN strækker sig ud over aktiveringen af KEAP1-Nrf2-ARE-systemet og berettiger yderligere forespørgsler i betragtning af den nuværende brug af dette middel i flere kliniske forsøg.
Sulforaphane er en Nrf2-uafhængig hæmmer af mitokondriel fission
Sulforaphane (SFN) er en isothiocyanatforbindelse, der er afledt i kosten oftest fra korsblomstrede grøntsager [56]. Det genereres i planter som en xenobiotisk reaktion på prædation via vesikulær frigivelse af det hydrolytiske enzym myrosinase fra beskadigede celler; dette enzym omdanner glucosinolater til isothiocyanter [42]. I løbet af de sidste to årtier er SFN i vid udstrækning blevet karakteriseret for dets rapporterede anticancer-, antioxidant- og antimikrobielle egenskaber [57]. Meget af denne effektivitet er blevet tilskrevet SFN's evne til at modulere KEAP1-Nrf2-antioxidant respons element (ARE) signalvejen, selvom yderligere aktiviteter af forbindelsen er blevet identificeret, herunder hæmning af histon deacetylase aktivitet og cellecyklus progression [ 29]. Nrf2 er hovedantioxidant-transkriptionsfaktoren, og under betingelser med homeostase undertrykkes dens stabilitet gennem virkningen af det cytoplasmatiske Cullin3KEAP1 ubiquitin-ligasekompleks [20]. Specifikt rekrutteres Nrf2 til Cullin3KEAP1-ligasen ved at binde til den dimere substratadapter KEAP1 og modificeres efterfølgende med polyUb-kæder, der målretter transkriptionsfaktoren for proteasom-medieret nedbrydning. Denne konstitutive omsætning begrænser halveringstiden for Nrf2 i ustressede celler til ~15 minutter [30], [33], [46], [55]. Som reaktion på adskillige typer stress, især oxidativ stress, fungerer KEAP1, et cysteinrigt protein, som en redoxsensor, og oxidativ modifikation af kritiske cysteiner, især C151, af KEAP1 dissocierer Nrf2-KEAP1 fra CUL3 og forhindrer derved Nrf2-nedbrydning [ 8], [20], [55]. Navnlig efterligner SFN og muligvis andre Nrf2-aktivatorer oxidativt stress ved at modificere C151 af KEAP1 f.eks. [21]. Stabilisering af Nrf2 muliggør dets translokation til kernen, hvor det inducerer ekspressionen af et batteri af fase II-antioxidant- og afgiftningsgener. Nrf2 binder sig til antioxidantresponspromotorelementerne (ARE) af dets beslægtede målgener gennem heterodimerisering med små Maf-proteiner [19]. Dette system præsenterer en dynamisk og følsom reaktion på indirekte antioxidanter som SFN, frie radikaler genereret af mitokondrierne [16] eller andre fysiologiske kilder til oxidativt stress [41].
Mitokondrier er dynamiske, subcellulære organeller, der regulerer et væld af cellulære funktioner lige fra ATP-produktion og intracellulær calciumbuffring til redoxregulering og apoptose [13], [49]. Mitokondrier repræsenterer også den vigtigste kilde til reaktive oxygenarter (ROS) i cellen. Korrekt regulering af mitokondriefunktionen er derfor nødvendig for at optimere ATP-produktionen for at imødekomme cellulære behov og samtidig minimere de potentielt skadelige virkninger af overdreven frie radikalproduktion. Et kritisk krav til finmodulering af mitokondriefunktion er mitokondriers evne til at fungere både uafhængigt som biokemiske maskiner og som en del af et stort, responsivt netværk.
Mitokondrielt netværks morfologi og funktion er bestemt af en reguleret balance mellem fission og fusion. Mitokondriel fission er påkrævet for dattercellearvning af mitokondrier under celledeling [28] såvel som for selektiv, autofagisk nedbrydning af depolariserede eller beskadigede mitokondrier, kaldet mitofagi [1]. Omvendt er fusion påkrævet for komplementering af mitokondrielle genomer og deling af elektrontransportkædekomponenter mellem nabomitokondrier [54]. På molekylært niveau reguleres mitokondriel fission og fusion af store, dynaminlignende GTPaser. Tre enzymer regulerer primært fusion: Mitofusin 1 og 2 (Mfn1/2) er to-pass ydre membranproteiner, der medierer ydre membranfusion via heterotypiske interaktioner mellem tilstødende mitokondrier [15], [25], [37], mens OPA1 er en indre membran. membranprotein, der samtidig sikrer matrixforbindelse ved at regulere sammensmeltningen af indre membraner [5]. GTPase-aktiviteten af alle tre proteiner er påkrævet for robust fusion [5], [18], og OPA1 reguleres yderligere af kompleks proteolyse inde i mitokondrernes indre membran af proteaserne OMA1 [14], PARL [6] og YME1L [45 ]. Det er vigtigt, at intakt mitokondriemembranpotentiale er påkrævet for effektiv fusion for at undertrykke integration af beskadigede og sunde mitokondrier [26].
Mitokondriel fission er primært katalyseret af et cytosolisk protein kaldet Dynamin-relateret protein 1 (Drp1/DNM1L). Drp1 rekrutteres fra cytosolen til potentielle fissionssteder på den mitokondrielle ydre membran [43]. De vigtigste receptorer for Drp1 på den ydre membran er mitokondriel fissionsfaktor (Mff) [32] og i mindre grad Fission 1 (Fis1) [51]. Derudover blev en lokkereceptor, MIEF1/MiD51, opdaget, der virker til yderligere at begrænse aktiviteten af Drp1-protein på potentielle fissionssteder [58]. Når først den er forankret ved den ydre mitokondriske membran, oligomeriseres Drp1 til spirallignende strukturer rundt om mitokondriets krop og bruger derefter energien afledt af GTP-hydrolyse til at mediere den fysiske spaltning af de mitokondrielle ydre og indre membraner [17]. Endoplasmatiske retikulum-afledte tubuli fungerer som en indledende sammensnøring af mitokondrier før Drp1-oligomerisering, hvilket understreger åbenbaringen, at ikke-indsnævrede mitokondrier er bredere end den tilladelige omkreds af en fuldført Drp1-spiral [12]. Aktin-dynamik er også vigtig for ER-mitokondrier-interaktioner, der går forud for mitokondriel fission [24]. Ud over sin rolle i mitokondriel fission katalyserer Drp1 fissionen af peroxisomer [40].
Drp1 ligner meget det velkarakteriserede dynaminprotein, idet begge proteiner indeholder et N-terminalt GTPase-domæne, et mellemdomæne, der er kritisk for selv-oligomerisering, og et C-terminalt GTPase-effektordomæne [31]. Drp1 opnår selektivitet for mitokondrielle membraner gennem en kombination af interaktioner med dets receptorproteiner Mff og Fis1 og også gennem dets affinitet for mitokondriespecifikke phospholipid cardiolipin via det unikke B-insert-domæne af Drp1 [2]. Drp1 eksisterer typisk som en homotetramer i cytoplasmaet, og samling af højere orden ved mitokondrielle fissionssteder medieres af det midterste domæne af Drp1 [3].
I betragtning af den implicitte forbindelse mellem mitokondriel funktion og KEAP1-Nrf2-ARE-vejen, undersøgte vi virkningerne af Nrf2-aktivering på mitokondriel struktur og funktion. Vi demonstrerer her, at SFN inducerer mitokondriel hyperfusion, der uventet er uafhængig af både Nrf2 og KEAP1. Denne effekt af SFN er gennem en hæmning af Drp1-funktionen. Vi demonstrerer yderligere, at SFN giver resistens over for apoptose, der er Nrf2-uafhængig og efterligner, som observeres i celler, der er udtømt for Drp1. Disse data tilsammen indikerer, at ud over at stabilisere og aktivere Nrf2, modulerer SFN mitokondriel dynamik og bevarer cellulær fitness og overlevelse.
Resultater
Sulforaphane inducerer Nrf2/KEAP1-uafhængig hyperfusion af mitokondrier
I løbet af undersøgelsen af virkningerne af Nrf2-aktivering på mitokondriel netværksdynamik opdagede vi, at behandling af udødeliggjorte, humane retinale pigmentepitelceller (RPE-1) med sulforaphane (SFN), en potent aktivator af Nrf2-signalering, inducerede en robust fusion af mitokondrielle netværk sammenlignet med vehikel-behandlede kontrolceller (fig. 1A og B). Morfologien af mitokondrierne i disse celler lignede i høj grad mitokondriernes morfologi i celler udtømt af siRNA af endogen Drp1, den vigtigste mitokondrielle fissionsfaktor (fig. 1A). Dette resultat rejste den spændende idé om, at mitokondriel fission og fusionsstatus reagerer direkte på Nrf2-niveauer i cellen. Stimulering af celler med andre Nrf2-stabilisatorer og -aktivatorer såsom proteasomhæmmeren MG132, pro-oxidanten tBHQ eller knockdown af Nrf2-hæmmeren KEAP1 inducerede imidlertid ikke mitokondriel fusion (fig. 1A og B). Stabilisering af Nrf2 ved disse manipulationer blev bekræftet ved western blotting for endogen Nrf2 (fig. 1C). Ydermere var ekspression af Nrf2 uundværlig for SFN-induceret mitokondriel fusion, da knockdown af endogen Nrf2 med siRNA ikke kunne modvirke denne fænotype (fig. 1D�F). Fordi SFN stimulerer KEAP1-Nrf2-ARE-vejen ved kovalent at modificere cysteinrester af KEAP1 [21], slog vi KEAP1 ned for at adressere, om SFN-induceret mitokondriel hyperfusion stimuleres gennem en KEAP1-afhængig, men Nrf2-uafhængig vej. Men udtømning af KEAP1 kunne heller ikke ophæve SFN-induceret mitokondriel fusion (fig. 1G�I). Faktisk vendte SFN profissionsmorfologien induceret af udtømning af KEAP1 (fig. 1G, panel b versus panel d). Disse resultater indikerer, at SFN-behandling forårsager mitokondriel fusion uafhængig af den kanoniske KEAP1-Nrf2-ARE-vej og førte os til at undersøge, om SFN direkte påvirker komponenter i mitokondriel fission eller fusionsmaskineri.
Figur 1 SFN inducerer Nrf2/KEAP1-uafhængig mitokondriel fusion. (A) RPE-1-celler blev transficeret med de angivne siRNA'er og 3 dage senere behandlet med DMSO eller Nrf2-aktivatorerne SFN (50 μM), MG132 (10 μM) eller tBHQ (100 μM) i 4 timer. Mitokondrier (rød) er mærket med et anti-Tom20-antistof, og kerner (blå) modfarves med DAPI. (B) Graf, der viser kvantificering af mitokondriel morfologi scoring fra (A). >50 celler pr. tilstand blev evalueret på en blind måde. (C) Repræsentative western blots fra (A). (D) RPE-1-celler blev transficeret med 10 nM siRNA og 3 dage senere behandlet med SFN i 4 timer før de blev fikseret og farvet som i (A). (E) Graf, der viser kvantificering af mitokondriel fænotype scoring fra (D). >100 celler pr. tilstand blev evalueret på en blind måde. (F) Repræsentative western blots fra (D). (G) Celler blev transficeret og behandlet som i (D) med siCON eller siKEAP1. (H) Celler fra (G) blev bedømt som i (B) og (E) på basis af mitokondriel morfologi. (I) Repræsentative western blots fra (G). Data i (B), (E) og (H) blev kompileret fra 3 uafhængige eksperimenter hver, og statistisk signifikans blev bestemt ved to-halet Student's t-test. Fejlbjælker afspejler +/- SD (For fortolkning af referencerne til farve i denne figurforklaring henvises læseren til webversionen af denne artikel).
Sulforaphane hæmmer Mitochondrial Association of Drp1
Baseret på opdagelsen af, at SFN-behandling inducerer mitokondriel hyperfusion, begrundede vi, at denne fænotype enten var en konsekvens af overdreven fusionsaktivitet eller en hæmning af fissionsaktivitet. For at skelne mellem disse to muligheder sammenlignede vi morfologien af peroxisomer i nærvær og fravær af SFN. Peroxisomer ligner mitokondrier, idet de er dynamiske organeller, hvis form og længde konstant er i flux [44]. Peroxisomer indeholder både Fis1 og Mff i deres ydre membran og er som konsekvens mål for Drp1-medieret fission [22], [23]. Imidlertid udnytter peroxisomer ikke fusionsmaskineriet i mitokondrielle netværk og gennemgår derfor ikke fusion [39]. Snarere modvirkes peroxisomal fission af forlængelsen af eksisterende peroxisomer via de novo-tilsætning af membraner og proteiner [44]. Fordi peroxisomer mangler Mfn1/2 og OPA1, begrundede vi, at hvis SFN aktiverer fusionsmaskineriet i stedet for at hæmme fissionsmaskineriet, ville peroxisomlængden ikke blive påvirket. I vehikelbehandlede celler opretholdes peroxisomer som korte, runde, punktformede organeller (fig. 2, panel b og d). Imidlertid øgede SFN-behandling peroxisomlængden med ~2 gange sammenlignet med kontrolceller (fig. 2, paneler f og h). Ydermere blev mange af peroxisomerne klemt i nærheden af midten, hvilket indikerer en potentiel spaltningsdefekt (fig. 2, panel h, pilespidser). Ligeledes var peroxisomer i celler transficeret med Drp1 siRNA unormalt lange (fig. 2, paneler j og l), hvilket bekræfter, at Drp1 er påkrævet til peroxisomal fission og antyder, at SFN-behandling forårsager mitokondrielle og peroxisomale fænotyper ved at forstyrre fissionsmaskineriet.
Figur 2 SFN inducerer peroxisomal forlængelse. (A) RPE-1-celler blev transficeret med 10 nM af det angivne siRNA og 3 dage senere behandlet med DMSO eller 50 µM SFN i 4 timer. Peroxisomer (grønne) blev mærket med et anti-PMP70 antistof, mitokondrier med MitoTracker (rød) og DNA modfarvet med DAPI. Forstørrede indsættelser af peroxisomer er vist til højre (paneler d, h og l) for at lette visualisering af ændringerne i morfologi induceret af SFN- og Drp1-udtømning. Pilespidser fremhæver indsnævringspunkter. (For fortolkning af referencerne til farve i denne figurforklaring henvises læseren til webversionen af denne artikel).
Vi bestemte derefter, hvordan SFN begrænser Drp1-funktionen. Mulighederne inkluderede reduktioner i ekspressionsniveauer, rekruttering/retention ved mitokondrier, oligomerisering eller enzymatisk aktivitet af GTPase. Et underskud i en af disse ville resultere i reduceret mitokondriel fission og hyperfusion. Vi påviste ikke reproducerbare ændringer i Drp1-proteinniveauer efter SFN-behandling (fig. 1C og 3A), og konkluderede derfor, at SFN ikke ændrer Drp1-stabilitet eller ekspression, hvilket stemmer overens med Drp1, der har en halveringstid på >10 timer [50] og vores SFN-behandlinger er af kortere varighed. Dernæst undersøgte vi, om SFN påvirkede rekrutteringen eller fastholdelsen af Drp1 til mitokondrier. Fraktioneringsundersøgelser viste, at SFN inducerede et tab af Drp1 fra mitokondriefraktionen (Fig. 3A, bane 7-8 og Fig. 3B). Som tidligere rapporteret [43] er kun en mindre brøkdel af Drp1 (~3%) forbundet med mitokondrielle netværk på et givet tidspunkt under steady state-betingelser med det meste af enzymet i cytoplasmaet (fig. 3A, bane 5-8) ). Disse fraktioneringsdata blev bekræftet ved hjælp af co-lokaliseringsanalyse, som viste en ~40% reduktion i mitokondrier-lokaliserede, punktformede Drp1-foci efter SFN-behandling (fig. 3C og D). Sammen indikerer disse data, at den mitokondrielle fusion induceret af SFN, i det mindste delvist, skyldes den svækkede association af Drp1 med mitokondrierne. Vores data skelner ikke mellem, om SFN interfererer med den mitokondrielle rekruttering versus den mitokondrielle tilbageholdelse af Drp1, eller begge dele, da analysen af endogen Drp1 ikke var egnet til at visualisere GTPase ved levende cellemikroskopi.
Figur 3 SFN forårsager et tab af Drp1 fra mitokondrierne. (A) Subcellulær fraktionering af RPE-1-celler efter 4 timers DMSO eller SFN. Helcellelysater (WCL), nukleare (Nuc), cytosoliske (Cyto) og rå mitokondrielle (Mito) fraktioner blev opløst ved SDS-PAGE og behandlet til western blotting med de angivne antistoffer. Migrationen af molekylvægtsmarkører er angivet til venstre. (B) Grafer, der viser densitometrisk kvantificering af Drp1 i de angivne fraktioner fra (A). (C) RPE-1-celler blev transficeret med 10 nM siCON eller siDrp1 og 3 dage senere behandlet med DMSO eller SFN i 4 timer. Drp1 (grøn) blev visualiseret med et anti-Drp1 antistof, mitokondrier med MitoTracker (rød) og kerner med DAPI (blå). (D) Automatiseret co-lokaliseringsanalyse af Drp1 og MitoTracker signal fra (C). Data i (B) og (D) blev kompileret fra henholdsvis 3 og 5 uafhængige eksperimenter, og statistisk signifikans blev bestemt ved to-halet Student's t-test. Fejlbjælker afspejler +/- SD og stjerner angiver statistisk signifikans. (For fortolkning af referencerne til farve i denne figurforklaring henvises læseren til webversionen af denne artikel).
Sulforaphane giver beskyttelse mod staurosportin-induceret apoptose uafhængig af Nrf2
Tidligere arbejde har vist, at mitokondriel fission er permissiv i dannelsen af porer i den ydre mitokondrielle membran genereret af Bax/Bak under apoptose [11]. Drp1 har vist sig at blive selektivt rekrutteret til mitokondrier under apoptose [11], og i overensstemmelse med dette er fragmenterede mitokondrier blevet observeret tidligt i processen [27]. Omvendt menes inhibering af mitokondriel fission at hæmme apoptose ved at blokere dannelsen af de ydre membranporer, der tillader cytokrom c-frigivelse [53]. Følgelig forsinker stimulering af mitokondriel fusion progressionen af apoptose induceret af forbindelser, herunder staurosporin (STS) [14]. For at bestemme, om SFN beskytter RPE-1-celler mod STS-medieret apoptose, og i givet fald, om dette kræver Nrf2, etablerede vi et assay til let at inducere poly ADP-ribosepolymerase (PARP) spaltning, et substrat af aktiveret caspase-3 og definitiv markør for apoptose. Behandling af RPE-1-celler med 1 ¿M STS i 6 timer forårsagede kun en meget beskeden spaltning af PARP, men dette blev forhindret af SFN-sambehandling (f.eks. Fig. 4A, bane 3 versus 4). For at øge robustheden af dette assay sensibiliserede vi yderligere celler til STS-induceret apoptose ved at forbehandle dem med siRNA rettet mod den anti-apoptotiske faktor, Bcl-XL. Denne forbehandling reducerede ekspressionen af Bcl-XL og fremmede markant PARP-spaltning som funktion af tid udsat for STS (fig. 4B, sammenlign bane 2 med bane 4-10). Det er vigtigt, at 2 timers forbehandling med SFN mildnet PARP-spaltning i celler udsat for STS (fig. 4C, bane 3 versus 4 og bane 5 versus 6). Ligeledes blev celler stabilt udtømt for Nrf2 af CRISPR/Cas9 sammenligneligt beskyttet mod STS-toksicitet ved SFN-forbehandling (fig. 4C, bane 11 versus 12 og bane 13 versus 14 og fig. 4D). Denne beskyttelse blev observeret under anvendelse af både PARP-spaltning (fig. 4C og D) og cellulær morfologi (fig. 4E) som udlæsninger. Effektiviteten af Nrf2-depletering ved CRISPR/Cas9 blev bekræftet ved western blotting (fig. 4C, Nrf2 blot). Som forudsagt blokerede udtømmende celler af Drp1, som også giver en hyperfusionsfænotype (fig. 1A), også PARP-spaltning som svar på STS sammenlignet med kontrolceller inkuberet med SFN (fig. 4F og G). Tilsammen er disse resultater i overensstemmelse med SFN, der giver beskyttelse mod apoptose gennem dets evne til at begrænse Drp1-funktionen, uafhængigt af stabiliseringen og aktiveringen af Nrf2.
Figur 4 De cytobeskyttende virkninger af SFN er uafhængige af Nrf2-ekspression (A) RPE-1-celler blev forbehandlet med DMSO eller 50 µM SFN i 2 timer før behandling med DMSO, 1 µM staurosporin (STS) eller 50 µM SFN. M etoposid i 6 timer og blev behandlet til anti-PARP western blotting. (B) RPE-1-celler blev transficeret med 2.5 nM siCON, 1 nM siBcl-XL eller 2.5 nM siBcl-XL og 3 dage senere blev behandlet med DMSO eller 1 μM STS i 2, 4 eller 6 timer. Repræsentative western blots er vist, og migrationen af molekylvægtsmarkører er angivet til venstre. (C) CRISPR/Cas9-genererede vildtype (Nrf2WT) og Nrf2 knockout (Nrf2KO) RPE-1-celler blev transficeret med 1 nM siBcl-XL og 3 dage senere blev forbehandlet med DMSO eller 50 μM SFN i 2 timer . Efterfølgende blev cellerne behandlet med 1 μM STS i 2, 4 eller 6 timer. Repræsentative western blots med de angivne antistoffer er vist. (D) Kvantificering af spaltet PARP som en procentdel af total PARP (spaltet+uspaltet) fra 3 uafhængige eksperimenter. Det er vigtigt, at niveauerne af spaltet PARP var sammenlignelige, uanset om celler udtrykte Nrf2 eller ej, hvilket indikerer, at SFN-beskyttelse fra STS er uafhængig af transkriptionsfaktoren. (E) 20X fase-kontrast billeder taget umiddelbart før høst af lysater fra (C). Målestok = 65 �m. (F) Repræsentative western blots, der viser, at udtømning af Drp1 giver næsten sammenlignelig beskyttelse mod STS som SFN-behandling. RPE-1-celler blev transficeret med 1 nM siBcl-XL og yderligere transficeret med enten 10 nM siCON eller 10 nM siDrp1. 3 dage senere blev siCON-celler forbehandlet med SFN som i (A) og (C) og derefter udsat for STS i 4 timer før de blev høstet og behandlet til western blotting med de angivne antistoffer. (G) Samme som (D) for dataene præsenteret i (F) kompileret fra 3 uafhængige eksperimenter. Fejlbjælker afspejler +/- SEM
Diskussion
Vi har opdaget, at SFN modulerer mitokondriel fission/fusionsdynamik uafhængigt af dets virkninger på KEAP1-Nrf2-ARE-vejen. Dette er spændende på grund af en formodet sammenhæng mellem mitokondriel dysfunktion og ROS-produktion og nødvendigheden af at dæmpe mitokondrier-afledte frie radikaler gennem aktiveringen af Nrf2. Denne yderligere funktionelle virkning af SFN er af potentiel betydning i betragtning af de mere end 30 kliniske forsøg, der i øjeblikket er i gang med at teste SFN til behandling af en række sygdomme, herunder prostatacancer, obstruktiv lungesygdom og seglcellesygdom [7], [10], [ 47].
Fordi SFN er et isothiocyanat [56] og det aktiverer Nrf2-signalering ved direkte at acylere kritiske KEAP1-cysteiner for at undertrykke Nrf2-nedbrydning [21], følger det, at SFN udøver sine pro-fusionseffekter ved at modulere aktiviteten af en fissions- eller fusionsfaktor via cysteinmodifikation . Vores data understøtter kraftigt, at Drp1 er negativt reguleret af SFN, selvom om GTPase er et direkte mål for acylering, mangler at blive belyst. På trods af dette vidensgab bliver funktionen af Drp1 klart kompromitteret af SFN, da både mitokondrier og peroxisomer bliver hyperfusionerede som svar på SFN-behandling, og disse organeller deler Drp1 for deres respektive spaltningshændelser [38]. Derudover reducerer SFN mængden af Drp1, der lokaliseres og akkumuleres ved mitokondrier (fig. 3). Fordi vores eksperimenter blev udført med alle endogene proteiner, er vores påvisning af Drp1 på mitokondrielle fissionssteder under steady-state betingelser, og vi kan derfor ikke skelne mellem en rekruttering versus en retentionsdefekt af enzymet forårsaget af SFN. Yderligere kan vi ikke eliminere muligheden for, at SFN acylerer en receptor ved mitokondrierne (Fis1 eller Mff) for at blokere Drp1-rekruttering endnu, vi formoder, at Drp1 er direkte modificeret. Drp1 har ni cysteiner, hvoraf otte ligger inden for det midterste domæne, der er påkrævet til oligomerisering [3], og en af dem ligger i GTPase Effector Domain (GED) ved C-terminalen af Drp1. Direkte acylering af nogen af disse cysteiner kan forårsage en aktivitetsdefekt i Drp1 og ligger derfor til grund for virkningen af SFN på mitokondriel dynamik. Især tyder tidligere arbejde på, at defekter i oligomerisering og katalytisk aktivitet kan ophæve tilbageholdelsen af Drp1 ved mitokondrierne [52]. Cys644 i GED-domænet er et særligt attraktivt mål baseret på tidligere arbejde, der viser, at mutation af dette cystein fænokopierer mutationer, der hæmmer Drp1 GTPase-aktivitet [4], og at dette særlige cystein modificeres af thiol-reaktive elektrofiler [9]. Løsning af dette udestående spørgsmål vil kræve massespektrometrisk validering. Sammenfattende har vi identificeret en ny, cytobeskyttende funktion for den klinisk relevante forbindelse SFN. Ud over at aktivere master-antioxidant-transkriptionsfaktoren Nrf2, fremmer SFN mitokondriel og peroxisomal fusion, og denne effekt er uafhængig af Nrf2. Mekanismen bag dette fænomen involverer en reduktion i funktionen af GTPase Drp1, den primære mediator af mitokondriel og peroxisomal fission. En væsentlig konsekvens af SFN-medieret mitokondriel fusion er, at celler bliver resistente over for de toksiske virkninger af apoptoseinduceren staurosporin. Denne yderligere cytobeskyttende virkning af SFN kunne være af særlig klinisk nytte i de talrige neurodegenerative sygdomme, for hvilke alder er den førende risikofaktor (f.eks. Parkinsons sygdom, Alzheimers sygdom, aldersrelateret makuladegeneration), da disse sygdomme er blevet forbundet med apoptose og reduceret niveauer og/eller dysregulering af Nrf2 [35], [36], [48].
Materialer og Metoder
Apoptoseanalyser
Celler blev podet og transficeret med siRNA som angivet nedenfor. Cellerne blev forbehandlet med 50 µM sulforaphan i 2 timer for at inducere mitokondriel fusion og blev derefter behandlet med 1 µM staurosporin for at inducere apoptose. På høsttidspunktet blev medier opsamlet i individuelle rør og underkastet højhastighedscentrifugering for at pelletere apoptotiske celler. Denne cellepellet blev kombineret med adhærente celler og solubiliseret i 2 gange koncentreret Laemmli-buffer. Prøver blev udsat for anti-PARP western blotting.
CRISPR/Cas9 Construct Generation
For at skabe LentiCRISPR/eCas9 1.1 blev LentiCRISPR v2 (addgene #52961) først skåret med Age1 og BamH1. Dernæst blev SpCas9 fra eSpCas9 1.1 (addgen #71814) PCR-amplificeret med Age1- og BamH1-overhæng under anvendelse af følgende primere (Forward AGCGCACCGGTTCTAGAGCGCTGCCACCATGGACTATAAGGACCACGAC, Reverse AAGCGCGGATCCCTTTTTCTTTTTGGCC-vektoren skåret over i vektoren) og ligeret. sgRNA-sekvenser blev bestemt ved at bruge Benchling.com. Parametre blev sat til at målrette den kodende sekvens med den højeste on-target og laveste off-target score. De følgende sekvenser (targeting sekvens understregede, hs sgNFE2L2 # 1 sense CACCGCGACGGAAAGAGTATGAGC, antisense AAACGCTCATACTCTTTCCGTCGC; hs sgNFE2L2 # 2 sense CACCGGTTTCTGACTGGATGTGCT, antisense AAACAGCACATCCAGTCAGAAACC; hs sgNFE2L2 # 3 sense CACCGGAGTAGTTGGCAGATCCAC, antisense AAACGTGGATCTGCCAACTACTCC) blev annealet og ligeret ind BsmB1 skåret LentiCRISPR / eCas9 1.1. Lentiviralt inficerede RPE-1-celler blev udvalgt med puromycin og opretholdt som en poolet population. Knockout blev bekræftet ved immunfluorescens og western blotting.
Cellekultur og transfektioner
Humane retinale pigmentepitelceller transformeret med telomerase (RPE-1) (ATCC) blev dyrket i Dulbecco's Modified Eagle Medium (DMEM) indeholdende 1 g/L glucose suppleret med penicillin, streptomycin, 1X ikke-essentiel aminosyrecocktail (Life Technologies), og 10 % føtalt bovint serum (Life Technologies). Til siRNA-transfektioner blev 30,000-35,000 celler/ml podet natten over. Celler modtog 10 nM siRNA fortyndet i serumfrit DMEM og kombineret med 0.3 % interferintransfektionsreagens (PolyPlus). Til apoptosesensibilisering modtog celler 1 nM Bcl-XL siRNA. Celler blev høstet 2-3 dage efter transfektion.
Kemikalier, antistoffer og siRNA Oligos
Antistoffer mod β-tubulin (Cell Signaling), β-tubulin (Sigma), Drp1 (BD Biosciences), KEAP1 (Proteintech), Lamin B1 (Abcam), PARP (Cell Signaling), PMP70 (Abcam) og Tom20 (BD Biosciences) ) blev anvendt ved 1:1000 fortyndinger til western blotting og til immunfluorescens. In-house blev anti-Nrf2 kaninantistof brugt ved 1:2000 til western blotting [34], [59]. Sulforaphan (Sigma) og staurosporin (Tocris) blev anvendt ved henholdsvis 50 µM og 1 µM. siRNA'er mod Drp1 (Dharmacon), Nrf2 (Dharmacon), KEAP1 (cellesignalering) og Bcl-XL (cellesignalering) blev brugt ved 10 nM, medmindre andet er angivet.
Immunfluorescens og in vivo-mærkning
Celler podet på 18 mm glasdækglas blev behandlet med vehikel eller lægemiddel, fikseret i 3.7 % formaldehyd og derefter permeabiliseret i 0.2 % Triton X-100/PBS på is i 10 min. Primære antistoffer blev inkuberet i 3% bovint serumalbumin (BSA) i PBS natten over ved 4 °C. Efter PBS-vaske blev celler inkuberet i 1 time i artsegnede, Alexa488- eller Alexa546-, konjugerede sekundære antistoffer (fortyndet 1:1000) og 0.1 µg/ml DAPI (Sigma) i 3% BSA/PBS. Mitokondrier blev visualiseret enten ved anti-Tom20-immunfluorescens eller ved at inkubere celler i 200 nM MitoTracker Red CMXRos (Molecular Probes, Inc.) i serumfrit DMEM i 30 minutter ved 37 �C før fiksering.
Mikroskopi og billedanalyse
Immunofluorescensprøver blev set på et LSM710 Confocal mikroskop (Carl Zeiss). Mikrofotografier blev taget ved hjælp af 63X eller 100X olienedsænkningsmål og billeder justeret og forbedret ved hjælp af Adobe Photoshop CS6. Co-lokaliseringsanalyse blev udført ved hjælp af Carl Zeiss LSM710 co-lokaliseringsfunktion med tærskler manuelt indstillet, mens de var blindet for identiteten af prøverne. Skalabjælker overalt, medmindre andet er angivet, er 10 �m. Mitokondriel morfologi blev vurderet ved blindet scoring. Hvis mitokondrierne i en celle blev opretholdt som multiple, runde, diskriminerende puncta, blev cellen bedømt som �fission�. Hvis individuelle mitokondrier ikke kunne skelnes, og hele mitokondrielle netværk virkede kontinuerligt, blev cellen bedømt som �fusion�. Alle andre celler, inklusive dem med klyngede mitokondrier, blev bedømt som �mellemliggende�.
Subcellulære fraktioneringer
RPE-1-celler blev dyrket til konfluens. Efter en PBS-vask blev celler underkastet centrifugering ved 600 �g i 10 minutter og resuspenderet i 600 µl isoleringsbuffer (210 mM mannitol, 70 mM saccharose, 5 mM MOPS, 1 mM EDTA pH 7.4+1 mM PMSF). Suspensionen blev lyseret 30 gange i en Dounce-homogenisator. En fraktion af homogenatet blev konserveret som et helcellelysat. Resten blev underkastet centrifugering ved 800 �g i 10 minutter for at pelletere kerner. Supernatanter blev udsat for centrifugering ved 1500�g i 10 minutter for at fjerne resterende kerner og ulyserede celler. Denne supernatant blev udsat for centrifugering ved 15,000 �g i 15 minutter for at pelletere mitokondrier. Supernatanten blev bevaret som �cytosolfraktionen�. Pelleten blev vasket forsigtigt med PBS og resuspenderet i isoleringsbuffer. Proteinkoncentrationen af hver fraktion blev målt ved bicinchoninsyre (BCA) assay, og ækvivalente mængder af protein blev opløst ved SDS-PAGE.
Western Blotting
Celler blev vasket i PBS og solubiliseret i 2 gange koncentreret Laemmli solubiliseringspuffer (100 mM Tris [pH 6.8], 2 % SDS, 0.008 % bromphenolblåt, 2 % 2-mercaptoethanol, 26.3 % glycerol og 0.001 % Pyrinin Y). Lysater blev kogt i 5 minutter før påfyldning på natriumdodecylsulfat (SDS) polyacrylamidgeler. Proteiner blev overført til nitrocellulosemembraner, og membranerne blev blokeret i 1 time i 5% mælk/TBST. Primære antistoffer blev fortyndet i 5 % mælk/TBST og inkuberet med blottet natten over ved 4 °C. Peberrodsperoxidase (HRP)-konjugerede sekundære antistoffer blev fortyndet i 5 % mælk/TBST. Blots blev behandlet med forbedret kemiluminescens, og densitometriske kvantificeringer blev udført ved hjælp af ImageJ-software.
Sulforaphane er et kemikalie fra isothiocyanatsamlingen af organo-svovlstoffer opnået fra korsblomstrede grøntsager, herunder broccoli, kål, blomkål, grønkål og collards, blandt andre. Sulforaphane produceres, når enzymet myrosinase omdanner glucoraphanin, et glucosinolat, til sulforaphane, også kendt som sulforaphane-glucosinolat. Broccolispirer og blomkål har den højeste koncentration af glucoraphanin eller forstadiet til sulforaphane. Forskningsundersøgelser har vist, at sulforaphane forbedrer den menneskelige krops antioxidantevner for at forhindre forskellige sundhedsproblemer. Dr. Alex Jimenez DC, CCST Insight
Sulforaphane og dens virkninger på kræft, dødelighed, aldring, hjerne og adfærd, hjertesygdomme og mere
Isothiocyanater er nogle af de vigtigste planteforbindelser, du kan få i din kost. I denne video gør jeg det mest omfattende tilfælde til dem, der nogensinde er blevet lavet. Kort opmærksomhed span? Spring til dit yndlingsemne ved at klikke på et af nedenstående tidspunkter. Fuld tidslinje nedenfor.
Nøglesektioner:
00: 01: 14 - Kræft og dødelighed
00: 19: 04 - Aging
00: 26: 30 - Hjerne og adfærd
00: 38: 06 - Final recap
00: 40: 27 - Dosis
Fuld tidslinje:
00: 00: 34 - Introduktion af sulforaphane, et vigtigt fokus i videoen.
00: 01: 14 - Cruciferous vegetabilsk forbrug og reduktion af allårsag dødelighed.
00: 02: 12 - Prostatacancerrisiko.
00: 02: 23 - Kræftrisiko.
00: 02: 34 - Lungekræft i rygere risiko.
00: 02: 48 - Brystkræftrisiko.
00: 03: 13 - Hypotetisk: Hvad hvis du allerede har kræft? (Interventionel)
00: 03: 35 - Plausibel mekanisme, der driver kræft- og dødelighedsassosierende data.
00: 04: 38 - Sulforaphane og kræft.
00: 05: 32 - Dyrebevis, der viser stærk virkning af broccoli-spireekstrakt på udviklingen af blæretumor hos rotter.
00: 06: 06 - Virkning af direkte tilsætning af sulforaphan hos patienter med prostatacancer.
00: 07: 09 - Bioakkumulering af isothiocyanatmetabolitter i egentligt brystvæv.
00: 08: 32 - Inhibering af brystcancerstamceller.
00: 08: 53 - Historie lektion: brassicas blev etableret som at have sundhedsegenskaber selv i det gamle Rom.
00: 09: 16 - Sulforaphans evne til at øge kræftfremkaldende udskillelse (benzen, acrolein).
00: 09: 51 - NRF2 som en genetisk switch via antioxidant responselementer.
00: 10: 10 - Hvordan NRF2 aktivering øger kræftfremkaldende udskillelse via glutathion-S-konjugater.
00: 10: 34 - Spiraler øger glutathion-S-transferase og reducerer DNA-beskadigelse.
00: 11: 20 - Broccoli-spire drikker øger benzenudskillelsen med 61%.
00: 13: 31 - Broccoli spirehomogenat øger antioxidant enzymer i den øvre luftvej.
00: 15: 45 - Cruciferous vegetabilsk forbrug og dødelighed i hjertesygdomme.
00: 16: 55 - Broccoli spire pulver forbedrer blodlipider og overordnet hjertesygdomsrisiko hos type 2 diabetikere.
00: 19: 04 - Begyndelse af aldringssektionen.
00: 19: 21 - Sulforaphane beriget kost forbedrer levetiden for biller fra 15 til 30% (under visse forhold).
00: 20: 34 - Betydningen af lav inflammation for lang levetid.
00: 22: 05 - Cruciferous grøntsager og broccoli spire pulver synes at reducere et bredt udvalg af inflammatoriske markører hos mennesker.
00: 24: 14 - Musestudier tyder på, at sulforaphan kan forbedre adaptiv immunfunktion i alderdommen.
00: 25: 18 - Sulforaphane forbedrede hårvækst i en musmodel af skaldethed. Billede på 00: 26: 10.
00: 26: 30 - Begyndelse af hjerne- og adfærdssektionen.
00: 27: 18 - Effekt af broccoli-spireekstrakt på autisme.
00: 27: 48 - Virkning af glucoraphanin på skizofreni.
00: 28: 17 - Start af depression diskussion (sandsynlig mekanisme og undersøgelser).
00: 31: 21 - Mus undersøgelse ved hjælp af 10 forskellige modeller af stress-induceret depression viser sulforaphan på samme måde som fluoxetin (prozac).
00: 32: 00 - Undersøgelse viser direkte indtagelse af glucoraphanin hos mus er tilsvarende effektiv til forebyggelse af depression fra social nederlagsspændingsmodel.
00: 33: 01 - Begyndelsen af neurodegenerationsafsnittet.
00: 33: 30 - Sulforaphane og Alzheimers sygdom.
00: 33: 44 - Sulforaphane og Parkinsons sygdom.
00: 33: 51 - Sulforaphane og Hungtington's sygdom.
00: 35: 55 - Sulforaphane og neuronal plasticitet.
00: 36: 32 - Sulforaphane forbedrer læring som model af type II diabetes hos mus.
00: 37: 19 - Sulforaphane og duchenne muskeldystrofi.
00: 37: 44 - Myostatininhibering i muskel-satellitceller (in vitro).
00: 38: 06 - Late video recap: dødelighed og kræft, DNA-beskadigelse, oxidativ stress og betændelse, udslip af benzen, kardiovaskulær sygdom, type II diabetes, hjernens virkninger (depression, autisme, skizofreni, neurodegeneration), NRF2-pathway.
00: 40: 27 - Tanker om at finde ud af en dosis broccolispirer eller sulforaphane.
00: 41: 01 - Anecdoter på spiring hjemme.
00: 43: 14 - På tilberedningstemperaturer og sulforafanaktivitet.
00: 43: 45 - Gut-bakteriekonvertering af sulforaphan fra glucoraphanin.
00: 44: 24 - Tilskud fungerer bedre, når de kombineres med aktiv myrosinase fra grøntsager.
00: 44: 56 - Madlavningsteknikker og cruciferous grøntsager.
Opvarmning reducerer epithiospecificerende proteinaktivitet og øger sulforafandannelse i broccoli
Abstrakt
Sulforaphane, et isothiocyanat fra broccoli, er en af de mest potente fødevareafledte anticarcinogener. Denne forbindelse er ikke til stede i den intakte grøntsag, men dannes snarere af dens glucosinolat-precursor, glucoraphanin, ved virkningen af myrosinase, et thioglucosidase-enzym, når broccolivæv knuses eller tygges. Imidlertid har en række undersøgelser vist, at sulforaphanudbyttet fra glucoraphanin er lavt, og at en ikke-bioaktiv nitrilanalog, sulforaphanitril, er det primære hydrolyseprodukt, når plantevæv knuses ved stuetemperatur. Nylige beviser tyder på, at i Arabidopsis styres nitrildannelse fra glucosinolater af et varmefølsomt protein, epithiospecifierprotein (ESP), en ikke-katalytisk cofaktor af myrosinase. Vores mål var at undersøge virkningerne af opvarmning af broccolibuketter og spirer på dannelsen af sulforaphane og sulforaphane nitril, for at bestemme, om broccoli indeholder ESP-aktivitet, og derefter at korrelere varmeafhængige ændringer i ESP-aktivitet, sulforaphan-indhold og bioaktivitet, som målt ved induktion af fase II afgiftningsenzym quinonreduktase (QR) i cellekultur. Opvarmning af friske broccolibuketter eller broccolispirer til 60 �C før homogenisering øgede samtidig dannelsen af sulforaphan og reduceret sulforaphannitrildannelsen. Et betydeligt tab af ESP-aktivitet sideløbende med faldet i dannelsen af sulforaphanitril. Opvarmning til 70 �C og derover mindskede dannelsen af begge produkter i broccolibuketter, men ikke i broccolispirer. Induktionen af QR i dyrkede musehepatom Hepa lclc7-celler gik parallelt med stigninger i sulforaphandannelse.
Forvarmning af broccolibuketter og spirer til 60 �C øgede den myrosinase-katalyserede dannelse af sulforaphane (SF) i vegetabilske vævsekstrakter betydeligt efter knusning. Dette var forbundet med fald i dannelsen af sulforaphanitril (SF Nitril) og aktiviteten af epithiospecificerende protein (ESP).
Som konklusion er sulforaphane et fytokemikalie, der findes i broccoli og andre korsblomstrede grøntsager. En ukontrolleret mængde oxidanter forårsaget af både interne og eksterne faktorer kan forårsage oxidativ stress i den menneskelige krop, som i sidste ende kan føre til en række sundhedsproblemer. Sulforaphane kan aktivere produktionen af Nrf2, en transkriptionsfaktor, der hjælper med at regulere beskyttende antioxidantmekanismer, der kontrollerer cellens reaktion på oxidanter. Omfanget af vores information er begrænset til kiropraktik og rygsøjlesundhedsproblemer. For at diskutere emnet, er du velkommen til at spørge Dr. Jimenez eller kontakte os på�915-850-0900 .
Rygsmerte er en af de mest udbredte årsager til handicap og ubesvarede arbejdsdage over hele verden. Rygsmerter tilskrives den næst hyppigste årsag til lægekontorbesøg, der kun er større end infektioner i de øvre luftveje. Ca. 80 procent af befolkningen vil opleve rygsmerter mindst én gang i hele deres liv. Rygsøjlen er en kompleks struktur, der består af knogler, led, ledbånd og muskler, blandt andet blødt væv. På grund af dette kan kvæstelser og / eller forværrede forhold, såsom herniated diske, kan i sidste ende føre til symptomer på rygsmerter. Sportsskader eller personskader er ofte den hyppigste årsag til rygsmerter, men nogle gange kan de enkleste bevægelser have smertefulde resultater. Heldigvis kan alternative behandlingsmuligheder, såsom kiropraktisk pleje, hjælpe lindring af rygsmerter ved brug af rygtilpasninger og manuelle manipulationer, der i sidste ende forbedrer smertelindring.
IFM's Find A Practitioner-værktøj er det største henvisningsnetværk inden for Functional Medicine, skabt for at hjælpe patienter med at finde Functional Medicine-praktikere overalt i verden. IFM Certified Practitioners er anført først i søgeresultaterne på grund af deres omfattende uddannelse i Functional Medicine